Культивирование, идентификация и очистка герпесвирусов
СОДЕРЖАНИЕ: Вирусы простого герпеса типов 1 и 2, пути заражения, воздействие на человека, методы наращивания, очистки и титрования. Цитомегаловирус человека: опасное воздействие, репликативный цикл, идентификация. Общая характеристика герпесвируса саймири.Курсовая работа
на тему:
Культивирование, идентификация и очистка герпесвирусов
Введение
В группу герпесвирусов входят, вероятно, более 70 вирусов, имеющих широкий круг хозяев. Характеристика и классификация вирусов этой труппы детально рассмотрены Мэтьюсом. Герпесвирусы подразделяют на три подсемейства: -герпесвирусы; -герпесвирусы; -герпесвирусы. Разделение на подсемейства основано на нескольких критериях, в частности на характере репликации вирусов, круге хозяев, белковом составе и структуре генома.
В этой работе мы обсудим культивирование, очистку и титрование герпесвирусов, взяв в качестве примера по одному представителю каждого из подсемейств, указанных выше. Особое внимание, однако, будет уделено вирусам простого герпеса – наиболее интенсивно изучаемым вирусам этой группы. Будут рассмотрены цитометаловирус человека и герпесвирус саймири.
1. Вирусы простого герпеса типов 1 и 2
Эти два вируса входят в а-подгруппу герпесвирусов. Они вызывают разнообразные заболевания человека, такие, как воспаление десен, стоматит, менингит, энцефалит и венерическую форму воспаления гениталий. Эти вирусы вызывают заболевания в ослабленных организмах. Некоторые факты указывают на связь вируса простого герпеса типа 2 с раком шейки матки, а вируса простого герпеса типа 1 с некоторыми другими злокачественными новообразованиями. Именно поэтому данным герпесвирусам, в особенности последние 10 лет, уделяют пристальное внимание. За последнее время были разработаны высокоэффективные методы наращивания, очистки и титрования этих вирусов. Методы изучения ВПГ-1 – и ВПГ-2 сходны между собой.
1.1 Получение заготовок вируса
1.1.1 Клетки
Широкий круг хозяев ВПГ-1 и ВПГ-2 позволяет получать заготовки вирусов на различных клетках. Тем не менее, выбор чувствительной культуры не прост из-за того, что выход вирусов в различных клеточных линиях широко варьирует. На практике для получения заготовок вируса часто используют клетки линии Нер-2 или ВПК с КЗ. Работы, проведенные с ВПГ-1 и ВПГ-2 в нашей лаборатории, показывают, что выход различных штаммов отличается примерно в 10 раз для данных клеточных линий. Хотя из этих двух линий клетки ВНК менее требовательны к условиям культивирования, клетки обеих линий достигают высокой плотности в роллерных сосудах как из стекла Winchester, так и из пластика. В сосудах такого большого объема особенно важно поддерживать оптимальный рН. При использовании бикарбонатного буфера культуральную среду необходимо насыщать С02 еще до стерилизации. В противном случае клетки следует выращивать в атмосфере с высоким содержанием С02 . Аппаратуру для одновременного культивирования большого количества роллерных сосудов можно купить или изготовить в мастерских. Например, на рис. lO.l сфотографирован аппарат, изготовленный из рамы Dexion, колес Meccano и моторов Parvalux в тропическом исполнении.
Клетки указанных выше линий способны расти в различных питательных средах. Обычно мы используем автоклавируемую среду Игла в модификации Глазго с 10% ТС. Для клеток линии ВНК в среду следует добавить 10% триптозофосфатного бульона. Окончательный выбор клеточной линии зависит от ряда факторов, в частности от характера эксперимента. Естественно, что используемые для приготовления вирусных заготовок клеточные линии должны быть предварительно проверены на отсутствие микоплазмы.
1.1.2 Методы заражения
Основной метод заражения описан Ватсоном и др. Целесообразно перед началом работы получить первичную и вторичную заготовки вируса, которые по возможности следует разместить в нескольких холодильниках на –70°С. Как правило, рабочие заготовки получают из вторичных и таким образом исключают длительное пассирование вируса. При этом сводится к минимуму возможность получения вирусных заготовок, сильно отличающихся по номеру пассажа, что создает трудности в прямом сравнении данных, полученных в разных экспериментах. В большинстве случаев для получения вирусных заготовок мы используем роллерные культуры, почти достигшие монослоя. Во флаконах обычно содержится 5–108 клеток ВНК или 3-108 клеток Нер-2. Высокая концентрация клеток позволяет получить и высокий титр вирусной заготовки. Клеточный монослой промывают для удаления слабо прикрепленных клеток и затем из вторичной заготовки вносят разведенный вирус. Чаще всего мы предпочитаем заражать клетки при множественности инфекции 0,001–0,01 БОЕ/кл, что предотвращает накопление дефектных вирионов.

1.1.3 Наращивание вируса
Любая стандартная среда для культивирования клеток может быть использована для получения вирусных заготовок. Обычно для выращивания клеток мы используем среду с 10% ТС. Уменьшение концентрации сыворотки приводит к некоторому снижению выхода вируса, однако при этом экономится дорогостоящая сыворотка. Заготовки вируса получают, используя следующую методику:
1. Вирус адсорбируют 1 ч в небольшом объеме среды, достаточном для покрытия монослоя. Множественность инфекции при этом составляет 0,001–0,01 БОЕ/кл.
2. Добавляют необходимое количество среды для культивирования клеток.
3. Клетки инкубируют 2–3 дня при соответствующей температуре. Однако продолжительное наращивание вируса при низкой температуре может привести к получению материала, содержащего большой процент температур-но-чувствительных мутантов или по крайней мере вируса, лучше растущего при 32°С, чем при 37°С. По этой причине мы предпочитаем инкубацию при 37°С.
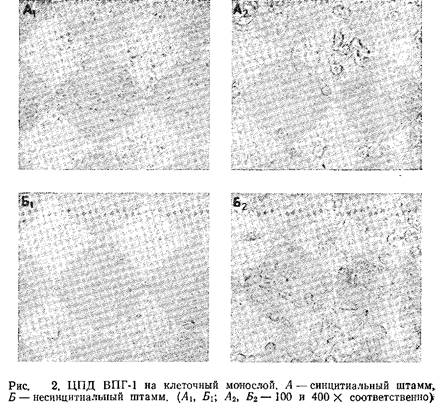
4. Через 2 дня инкубации при 37°С начинают появляться признаки ЦПД, однако сроки их появления могут варьировать в зависимости от штамма вируса и линии клеток. Как правило, ВПГ-2 размножается заметно быстрее, чем ВПГ-1. Как известно, существует два вида ЦПД: синцитиальное и несинцитиальное. Морфологическая картина ЦПД, вызванного этими двумя штаммами, показана на рис. 2.
5. Клетки собирают после максимального проявления признаков ЦПД. Методика сбора клеток зависит от штамма вируса и линии клеток. Так, в случае штамма HFEM и клеток линии ВНК основную массу среды можно удалить из культурального сосуда без потери вируса. Затем клетки собирают в оставшуюся среду стерильной резиновой палочкой и центрифугируют при малой скорости. При работе с другими штаммами вируса, например штаммом 186, инфицированные клетки удается собрать только центрифугированием культуральной среды, поскольку большая часть клеток отрывается от поверхности сосуда. Если лизис клеток уже произошел, необходимо использовать среду как составную часть заготовки вируса. Клетки собирают центрифугированием, а затем ресуспендируют в выбранной среде, например стерильной дистиллированной воде или же обычной культуральной среде. Заготовки можно ресуспендировать в среде с криопротектором, хотя это и не является строго обязательным.
6. Одной из серьезных проблем, возникающих по мере получения заготовок вирусов герпеса, является их очистка от клеточного материала, поскольку большая часть вируса тесно связана с клетками. Клетки разрушают, проводя ряд циклов замораживания и оттаивания, однако это может привести и к снижению титра вируса. Обработка клеток ультразвуком небезопасна из-за возможного образования аэрозолей, содержащих инфекционный вирус. Поэтому лучше всего проводить дезинтеграцию в ультразвуковой бане в закрытом сосуде. К сожалению, мы не можем рекомендовать какую-либо определенную марку ультразвукового дезинтегратора. Необходимо проверять эффективность каждого из них. Вирусные заготовки обрабатывают ультразвуком до полного разрушения клеток. Процедуру проводят в ледяной воде, чтобы не допустить сильного нагревания.
7. Клеточный дебрис удаляют центрифугированием, готовые заготовки разливают в ампулы и хранят, как описано ниже. Заготовки вируса герпеса длительное время лучше всего хранить при –70°С. Ни при каких обстоятельствах не следует хранить вирусы герпеса при –20°С, поскольку это приводит к значительному снижению инфекционного титра. В течение нескольких дней штамм HFEM можно хранить при 4°С, однако этот срок варьирует для разных штаммов. Оттаивание вируса необходимо проводить быстро, по возможности следует избегать повторного замораживания и оттаивания. Тем не менее, мы показали, что штамм HFEM, имеющий высокий титр в культуральной среде, теряет всего лишь 0,51og инфекционного титра при пятикратном замораживании и оттаивании. Мы обнаружили, что и при быстром оттаивании вирусных заготовок, хранящихся при –70°С даже в течение 5 лет, не происходит снижения титра вируса. Перед использованием каждую заготовку необходимо проверить на стерильность как на твердой, так и в обогащенной жидкой бактериальных средах. Лучше всего проверять заготовки под электронным микроскопом, чтобы исключить использование тех заготовок, в которых велико соотношение физических и инфекционных частиц. Мы установили, что с помощью описанных выше методов удается получать заготовки с титрами около 109 –1010 БОЕ/мл и низкими соотношениями физических и инфекционных частиц.
1.2 Электронная микроскопия
Подсчет частиц по методу 1 оор drop предложен Ватсоном. Вирус разводят в воде или в растворе, содержащем 0,2 мг ВСА/мл в случае вирусных препаратов с концентрацией белка ниже 0,2 мг/мл, В любом случае вирус разводят до концентрации 109 –1010 частиц/мл. Суспензию смешивают с равным объемом калибровочного материала. Мы рекомендуем использовать полистироловый латекс DOW, содержащий -109 латексных частиц/мл. К полученной смеси добавляют 0,12 мл нейтрального 2%-ного раствора фосфовольфрамата натрия. Одну каплю полученного материала наносят, а затем высушивают на формваровых сетках с ячейками размером: 400 меш. Сетки просматривают под электронным микроскопом при увеличении 40 000. Частицы, видимые под микроскопом, классифицируют как голые, покрытые оболочкой и покрытые оболочкой. Эти частицы представлены на рис. 5. Непроницаемые, покрытые оболочкой частицы выглядят как плотные белые шарики, однако можно показать их способность количественно превращаться в проницаемые, покрытые оболочкой частицы. Как правило, для характеристики каждого образца вируса просчитывают пять групп из 20 латексных частиц.
1.3 Определение инфекционного титра
Вирусы простого герпеса относительно легко титруются различными методами. Мы обычно используем суспензионный метод либо метод монослойных культур.
1.3.1 Титрование в суспензионной культуре
1. Готовят разведения вируса на культуральной среде. К2 мл каждого разведения добавляют 8-106 клеток ВНК.
2. Суспензию при постоянном перемешивании инкубируют 30 мин при 37°С. Затем добавляют 8 мл 2%-ной КМЦ в культуральной среде и перемешивают с вирусом и клетками. Для данного метода содержание ТС в среде может составлять не более 5%.
3. Приготовленные суспензии разливают на две чашки Петри диаметром 60 мм и инкубируют 2–3 дня при 37°С в атмосфере 5%-ного С02 .
4. Монослой клеток фиксируют при помощи обычных фиксаторов и окрашивают генциан-виолетом или метиленовым сидим.
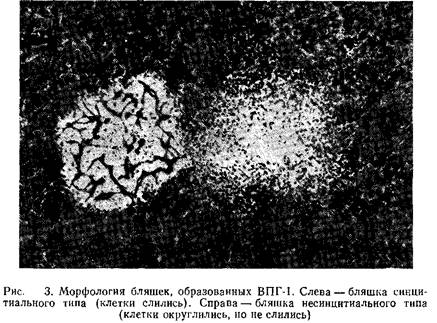
Вирусные бляшки можно обнаружить и невооруженным глазом, однако для их подсчета следует воспользоваться стереомикроскопом. Бляшки вирусов простого герпеса различаются по морфологии. Некоторые штаммы вызывают слияние клеток, что приводит к образованию больших синцитиальных бляшек, в то время как другие штаммы вызывают округление клеток. На рис. 3 для сравнения показана морфология описанных выше бляшек. Однако во многих случаях продолжительная инкубация клеток с вирусом, вызывающим образование синцитиев, приводит к изменению картины ЦПД. Бляшки при этом приобретают несинцитиальный характер.
1.3.2 Титрование на монослойной культуре
При использовании данного метода особенно важен выбор соответствующей клеточной линии. Как правило, инфицирование монослоя клеток ВНК приводит к образованию мелких бляшек. Кроме того, частота их возникновения, как свидетельствует наш опыт, также небольшая. При титровании вируса на монослойных культурах мы обычно используем клетки Vero, на которых эффективность бляшкообразования такая же, а для некоторых штаммов даже выше, чем на ВНК, находящихся в суспензии.
1. Клетки Vero рассевают в количестве 2–106 клеток на чашку за 18–24 ч до заражения.
2. Из чашек отсасывают среду и клетки инфицируют 0,1 мл соответствующего разведения вируса. Для адсорбции вируса чашки инкубируют 1 ч при 37°С.
3. В каждую чашку добавляют по 5 мл 2%-ной КМЦ, приготовленной на культуральной среде, и культуры инкубируют при 37°С. Время инкубации сильно варьирует в зависимости от штамма вируса. Быстрорастущий ВПГ-2 формирует хорошо различимые бляшки всего за 2 дня, в случае других вирусов требуется 4 дня.
4. Клетки фиксируют и окрашивают, как описано выше.
1.3.3 Среды для титрования
Существует множество разнообразных сред, используемых при титровании суспензионных и монослойных культур. Тем не менее мы предпочитаем среду, содержащую КМЦ, поскольку она удобна в работе и не ограничивает рост бляшек. Однако во время культивирования чашки должны находиться на ровной поверхности, и работать с ними следует осторожно, поскольку любое повреждение слоя КМЦ может привести к искажению картины бляшек.
1.3.4 Метод оспин
ВПГ-1 и ВПГ-2 формируют оспины на хорионаллантоисной мембране куриных эмбрионов. Существует предположение, что с помощью этого метода можно различить ВПГ-1 и ВПГ-2.
1.4 Приготовление экстрактов клеток, инфицированных вирусом
Тотальные гомогенаты клеток, инфицированных вирусом, или детергентные экстракты таких клеток обычно используются в качестве антигенов в различных иммунологических реакциях, таких, как ELISA и RIA. Кроме того, их применяют для иммунизации при получении гипериммунных антисывороток или моноклональных антител к белкам ВПГ. Поэтому выбор инфицируемой клеточной линии зависит от дальнейшего использования антигена.
1. Для радиоиммунологических методов используют клетки Vero или ВНК, которые заражают при множественности инфекции 10 БОЕ/мл.
2. Клетки вместе с культуральной средой собирают через 18–24 ч после заражения и центрифугируют при малой скорости.
3. Клеточный осадок дважды промывают PBS, а затем ресуспендируют в дистиллированной воде таким образом, чтобы концентрация клеток по возможности составляла 1-108 кл/мл.
4. Суспензию инфицированных клеток разрушают в ультразвуковой бане и хранят при –70°С. На рис. 6 показаны полипептиды, присутствующие в такой фракции.
Для некоторых методов, например иммунопреципитации, требуется фракция растворимого антигена. Такую фракцию получают центрифугированием суспензии разрушенных инфицированных клеток. С целью увеличения количества мембранных белков в этой фракции можно использовать детергенты. Свойства полученного антигена необходимо проверить при помощи ELISA или диффузией в геле по методу Оухтерлони, используя кроличью гипериммунную сыворотку к зараженным вирусом герпеса клеткам. Антиген для иммунизации получают, как описано выше, однако выбору клеток уделяют особое внимание. Например, для иммунизации животных с целью получения моноклональных антител используют антиген, выделенный из инфицированных L-клеток мыши, в то время как для получения кроличьих сывороток – антиген, выделенный из инфицированных клеток почки кролика RK13, растущих в присутствии 5% кроличьей сыворотки.
1.5 Получение очищенного вируса
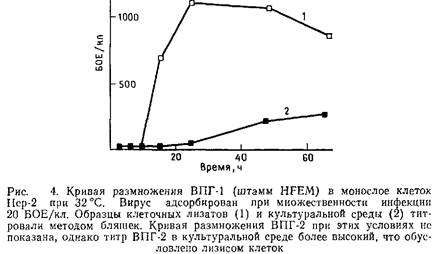
Методы выделения вирусов простого герпеса довольно сложны по сравнению с другими вирусами. Более того, метод, хорошо зарекомендовавший себя при работе с одним штаммом вируса на определенной клеточной линии, может оказаться неприемлемым для других штаммов на той же линии или того же штамма вируса на других клетках. Поэтому мы опишем два метода. Мы показали, что первый метод можно успешно применять для получения штамма HFEM и некоторых других при условии, что для выращивания вируса используют клетки Нер-2. Данный метод прост и позволяет получать вирусные препараты высокой степени чистоты. Второй метод позволяет достигнуть удовлетворительных результатов со всеми штаммами вируса и различными типами клеток. Для выбора оптимальных условий получения высокого выхода вируса и качественного исходного материала для дальнейшей очистки следует вначале изучить кривую роста исследуемого вирусного штамма на различных линиях клеток. На рис. 4 показана кривая роста штамма HFEM на клетках Нер-2 при высокой множественности инфекции и культивировании клеток при 32°С.
1.5.1 Метод 1
1. Культуры Нер-2 в состоянии монослоя заражают при множественности инфекции 20–25 БОЕ/кл.
2. После адсорбции вируса зараженные клетки промывают, добавляют свежую среду и культивируют 2–3 дня при 32°С.
3. При необходимости радиоактивного мечения вирусоспецифических белков в культуральную среду вносят изотопы через 4 ч после начала культивирования.
4. По окончании культивирования культуральную среду центрифугируют при низкой скорости для удаления клеточного дебриса.
5. Вирус осаждают ПЭГ в присутствии 0,5 М NaCl или центрифугированием в роторе GSA в центрифуге SorvallRCS-5B.
6. Вирус ресуспендируют в трис-буфере низкой молярности, рН 7,8, содержащем 50 мМ NaCl. Полученную суспензию по возможности лучше оставить на ночь.
7. Вирусную суспензию наслаивают на 30 мл 5–45%-ного градиента сахарозы в указанном выше буфере и центрифугируют 1 ч при 12500 об/мин. После центрифугирования в центре градиента четко видна рыхлая белая полоса очищенного вируса. Эта видимая невооруженным глазом полоса и есть единственная в градиенте фракция инфекционного вируса.
Полученный вирус уже достаточно очищен, и его можно использовать в различных целях, в частности для выделения вирусной ДНК. Из фракции вирус легко осадить обычным центрифугированием. Для получения вирусных препаратов высокой степени чистоты может быть использован ряд других методов, включая повторное центрифугирование в градиенте сахарозы или CsCl. Эти методы позволяют получать препараты вируса, в которых отношение белка к числу частиц составляет менее чем 20 мкг на 1010 частиц.
1.5.2 Метод 2
Данный метод был разработан Спир и Ройзманом и Хейном и др.
1. Заражение клеток проводят по методу 1.
2. Клетки инкубируют 18–24 ч при 37°С, затем их снимают с подложки и центрифугируют при малой скорости.
3. Полученный таким образом клеточный осадок ресуспен-дируют в стандартном буфере для ретикулоцитов и в течение 10 мин дают клеткам набухнуть. Цитоплазматическую фракцию клеток получают в гомогенизаторе Даунса.
4. Клеточный гомогенат центрифугируют при малой скорости для удаления ядер и клеточного дебриса.
5. Супернатант, содержащий большую часть инфекционного вируса, наслаивают на 5–40%-ный градиент декстрана, приготовленный на трис-буфере, и центрифугируют 1 ч при 12500 об/мин. Фракция вируса видна невооруженным глазом в центре градиента.
6. Вирус из фракции осаждают центрифугированием в течение 1 ч при 20000 об/мин.
Полученный вирус уже достаточно очищен и может быть использован в различных целях. Кроме того, степень чистоты вируса можно повысить, как описано выше. Очищенный вирус, полученный любым методом, ресуспендируют в дистиллированной воде или в соответствующем буфере. Аликвоты суспензии можно использовать для заражения клеток, подсчета общего числа частиц и определения концентрации белка. Вирус хранят при –70°С, однако следует учитывать, что оттаивание ведет к частичному разрушению вирусной оболочки. На рис. 5 показаны различные вирусные частицы, обнаруженные в препаратах очищенного вируса.
1.5.3 Анализ структурных вирусных полипептидов
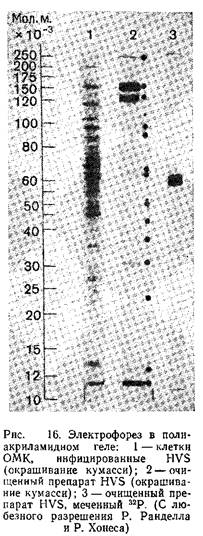
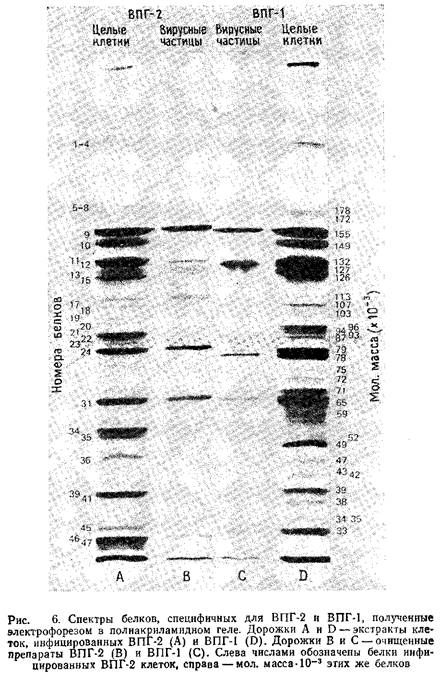
В большинстве случаев проводится электрофорез в пластинах полиакриламидного геля в присутствии ДДС-Na, как было описано впервые Даймэком и Ватсоном. Модификация данного метода изложена в работе. Гели окрашивают кумасси бриллиантовым синим, а для авторадиографического анализа экспонируют с рентгеновской пленкой. На рис. 6 представлены спектры структурных полипептидов ВПГ-1 и ВПГ-2, полученные гель-электрофорезом. Другие характеристики этих полипептидов приведены в табл. 1.
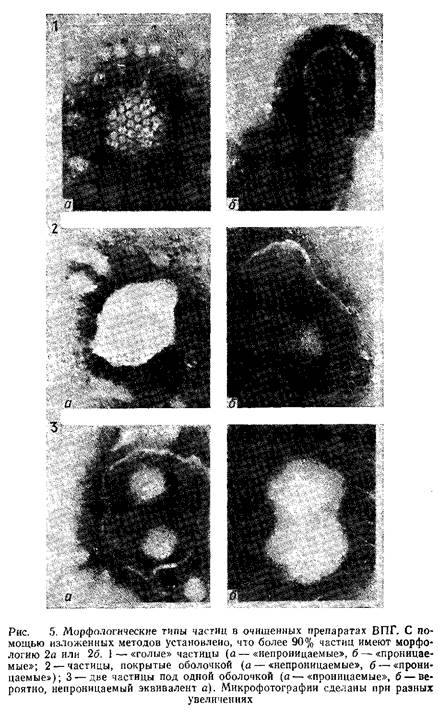
Таблица 1. Свойства и номенклатура некоторых белков ВПГ-1 и ВПГ-2
| ВПГ-2 | Свойства | ВПГ-1 | ||||
| номер белка | структурный вирусный белок | молекулярная масса | молекулярная масса | структурный вирусный белок | номер белка | |
ICSP 5–8 ICSP 9 ICSP 10 ICSP 11 ICSP 12 ICSP 13–15 ICSP 22 NN ISP 34–35 NN NN ICSP 46–47 |
NS VP5 NS NS VP7–8,5 NS NN NS NS NN VP22–23 |
182–186 157 153 146 143 126–140 85–90 92 54 42 4 53 28–30 |
Предранний фос-фопротеин Основной белок капсида Рибонуклеотидре-дуктаза Основной ДНК-связывающий белок Гликопротеины gA, gB Щелочная нуклеа-за Гликопротеин gE Белок, ассоциированный с ДНК-полимеразой Тимидинкиназа Гликопротеин gD, ответственный за перекрестную нейтрализацию между ВПГ-2 и ВПГ-1 Белок капсида |
175 155 149 132 112–126 85–90 80 48 44 59 30–33 |
NS VP5 NS NS VP7–8,5 NS NN NS NS VP18 VP22–23 |
ICP 4 ICP 5 ICP 6 ICP 8 ICP 9–14 ICP 19 NN ICP 29 NN ICP 31 ICP J39–40 |
1.6 Другие а-герпесвирусы
В нашей лаборатории выращены и очищены и многие другие а-герпесвирусы, включая: вирус мамиллита крупного рогатого скота; вирус псевдобешенства; вирус герпеса лошадей типа 1; вирус ринотрахеита кошек; вирус кошачьего сомика. С этими вирусами работают практически так же, как и с вирусами простого герпеса. Поэтому ниже мы отметим только особенности работы с каждым из перечисленных выше вирусов.
1.6.1 Вирус мамиллита крупного рогатого скота и вирус псевдобешенства
Эти вирусы дают на клетках ВНК высокие титры, которые легко определить суспензионным методом, предложенным Расселом. Для очистки вирусов клетки ВНК заражают с высокой множественностью инфекции и культивируют при 37°С 24 ч или 48 ч. На этой стадии вирусы могут быть очищены. Таким образом, получают качественные препараты оболочечных вирусов, белковый спектр которых представлен на рис. 7.
1.6.2 Вирус герпеса лошадей типа 1
Рестрикционный анализ показывает, что разные штаммы EHV-1 могут принадлежать как субтипу 1, так и субтипу 2. Первый связан с выкидышами у кобыл, хотя выделяют его и из респираторного тракта. Второй относится к вирусам респираторного типа и не связан с выкидышами. Вирусы субтипа 1 размножаются во многих культурах. В нашей лаборатории мы обычно используем клетки линии RK13, хотя в других лабораториях предпочитают использовать клетки лошадей. Кроме того, вирусы субтипа 1 успешно размножаются в культурах клеток L-M. Некоторые штаммы субтипа 1 были адаптированы к росту на хомяках. Вирусные изоляты субтипа 2 растут только в клетках лошадей. Используя метод 1, можно проводить очистку вирусов обоих субтипов. На рис. 7 показаны спектры полипептидов EHV-1, BMV, PRV, ВПГ-1 и ВПГ-2.
1.6.3 Вирус ринотрахеита кошек
Этот вирус специфичен для клеток кошачьего происхождения. Культивируют его так же, как и ВПГ-1. Для наращивания вируса мы используем, как правило, линию клеток кошки, представленную д-ром П. Тэлботом, однако можно использовать эмбриональные клетки легкого кошки, полученные доктором О. Джэр-ретом и поставляемые фирмой FlowLaboratories, Irvine, Scotland. Доктор P. Мэйес в градиенте тартрата калия успешно выделил вирус из культуральной среды инфицированных клеток почки кошек Crandell-Rees.
1.6.4 Вирус кошачьего сомика
Данный вирус имеет ограниченный круг хозяев и лучше всего размножается в клетках американского сомика-кошки. Методы культивирования и очистки этого вируса такие же, как и для ВПГ-1, за исключением того, что ВВ-клетки и вирус культивируют при 28°С.
2. Цитомегаловирус человека
Р-субгруппа герпесвирусов включает в себя цитомегаловирусы различных животных. Цитомегаловирусы человека вызывают различные заболевания, особенно у людей с ослабленной иммунной системой, а также в период беременности. Инфекция этими вирусами довольно часто приводит к смертельному исходу либо вызывает сильнейшее истощение больных с естественной или искусственной иммунной супрессией. Во время беременности этот вирус может вызвать уродства плода, выкидыши и повышенную чувствительность новорожденных к тяжелым инфекциям после родов. Именно по этим причинам в последние 10 лет резко возросла интенсивность изучения цитомегаловируса. Тем не менее, в этой области в отличие от вирусов простого герпеса не достигнуто значительного прогресса.
2.1 Получение заготовок вируса
Репликативный цикл цитомегаловируса человека продолжительнее, чем штаммов ВПГ. Кроме того, данный вирус прочно ассоциирован с клеткой и имеет высокую клеточную специфичность. Хотя во многих лабораториях для наращивания этого вируса используют клетки MRC-5, Flow 5000 или Нер-2, В. Гибсон рекомендует ранние пассажи на фибробластах крайней плоти человека или клетках легкого эмбриона человека. Гибсон культивирует перечисленные клетки в среде DMEM, содержащей 4500 мг/л глюкозы и 10% ЭТС. В нашей лаборатории используют клетки MRC-5, культивируемые в среде Игла в модификации Glasgow с 10% ЭТС. Заготовки вируса получают следующим образом:
1. Не дожидаясь образования плотного монослоя, добавляют CMV в минимальном объеме среды. Адсорбцию вируса проводят 1–2 ч при 37°С.
2. После адсорбции к клеткам добавляют необходимый объем культуральной среды и инкубируют их при 37°С.
3. Через 7–10 дней после заражения старую культуральную среду заменяют на свежую. Клетки продолжают инкубировать при 37°С до выявления признаков ЦПД.
4. Культуры ежедневно просматривают. Когда ЦПД достигнет своего максимума, клетки собирают в культуральную среду и центрифугируют 10 мин при 2000 об/мин. Собирают супернатант.
5. Осадок ресуспендируют в небольшом объеме культуральной среды, разрушают в ультразвуковой бане и хранят при –70°С.
6. Большая часть вируса содержится в культуральной среде, поэтому супернатант, полученный на стадии 4, центрифугируют 2 ч при 12000 об/мин в роторе GSA.
7. Полученный осадок ресуспендируют в небольшом объеме культуральной среды, осторожно разрушают в ультразвуковой бане и хранят при –70°С. Данный вирус не так стабилен при –70°С, как ВПГ. Поэтому его не следует подвергать частым замораживаниям и оттаиваниям. Оттаивание CMV, как и ВПГ, следует проводить быстро в теплой воде.
2.2 Цикл размножения
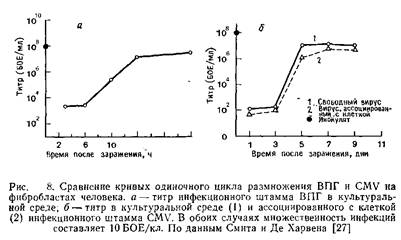
Смит и Де Харвен сравнили кривые роста ВПГ и CMV и определили время наступления тех или иных фаз инфекции. Результаты этих исследований приведены на рис. 8 и в табл. 2. Однако остается непонятным, насколько точно отражают данные кривые последовательность событий в зараженной клетке. Как уже отмечалось выше, цикл размножения CMV значительно длиннее, чем ВПГ.
Таблица 2. Время наступления основных этапов инфекции CMV и ВПГ. Сравнительные данные, полученные при помощи электронной микроскопии
| Этапы | Начало события | |
| CMV | ВПГ | |
| Слияние клеток и округление | 0,5 | 6 |
| Изменения аппарата Гольджи | 1 | 8 |
| Скрытый период | 2 | 2 |
| Появление большого количества гранул, напоминающих перихроматин | 2 | 3 |
| Конденсация хроматина | Не наблюдается | 3 |
| Сборка в ядрах первых капсидов | 3 | 4 |
| Появление первых капсидов, имею- | 3,5 | 5 |
| щих плотную сердцевину | ||
| Одевание ядерной мембраной | 3,5 | 6 |
| Появление голых капсидов в цито- | 4 | 6 |
| плазме | ||
| Появление плотных цитоплазматиче-ских агрегатов | 4 | Не наблюдается |
| Первые высвободившиеся из клетки частицы | 4 | 8 |
| Удвоение ядерной мембраны | 4 | 8 |
| Одевание цитоплазматической мембраной | 4 | 8 |
| Лизис клетки | 7–8 | 24–48 |
2.3 Идентификация вируса
Множество попыток было предпринято для разработки удобных методов титрования медленно растущих вирусов. В большинстве лабораторий остановились на методе определения конечной точки TCID50 . Сложность данной проблемы заключается в сохранении клеточного монослоя в течение времени, необходимого для формирования бляшек. В нашей лаборатории используют следующую модификацию метода бляшек, рекомендованную Вентвесом и Френчем:
1. Клетки линии MRC-5 высевают в 24-луночные планшеты или в чашки Петри диаметром 30 мм для культивирования клеток. Культуры, достигшие состояния монослоя, инфицируют вирусом, разведенным в 200 мкл культуральной среды.
2. По истечении первого часа адсорбции среду заменяют на 1%-ную агарозу, приготовленную на культуральной среде с 2% ЭТС. Клетки инкубируют при 37°С.
3. Через неделю еще раз добавляют необходимое количество агарозы и продолжают инкубацию при 37°С.
4. Через 2–3 нед с момента заражения под микроскопом удается обнаружить небольшие очаги инфицированных клеток. Незадолго до того, как разрушится монослой неинфицированных клеток, культуры фиксируют в забуференном формалине, осторожно удаляют агарозу и окрашивают 1%-ным генцианвиолетом. Небольшие очаги инфекции можно подсчитать под стереомикроскопом. Вместо генцианвиолета культуры можно окрашивать 1%-ным нейтральным красным. Кроме того, нам удалось получить бляшки CMV, используя КМЦ вместо агарозы.
2.4 Метод черных бляшек
Этот метод может быть использован для идентификации любых вирусов. Однако особенно удобен он оказался для CMV, образующего бляшки малого размера.
1. Культуры, достигшие монослоя, инфицируют обычным способом.
2. Через 6–7 дней после заражения удаляют КМЦ и клетки трижды промывают PBS. Затем в течение 5 мин клетки фиксируют 0,25%-ным раствором глутарового альдегида в PBS.
3. Отмыв фиксатор с помощью PBS, добавляют соответственно разведенную антисыворотку или моноклональные антитела против CMV. Клетки оставляют при комнатной температуре на 2 ч.
4. Клетки отмывают от антител PBS и инкубируют 3 ч при комнатной температуре со вторыми, обычно конъюгированными с пероксидазой хрена антителами против первых антител.
5. После трехкратной промывки в PBS в каждую культуру добавляют субстрат Hanker-Yates, 50 мл 0,1 M трис-HCl, pH 7,5, и 0,5 мл 1%-ной H2 02 ). Окраска проявляется в течение 5–30 мин.
6. Чашки промывают дистиллированной водой, переворачивают и дают им высохнуть в темноте. Темноокрашенные бляшки вируса идентифицируют и подсчитывают.
2.5 Культивирование и очистка CMV
1. Не дожидаясь образования плотного монослоя, культуры заражают вирусом.
2. По окончании адсорбции культуральную среду заменяют свежей, содержащей 2% ЭТС. Инфицированные клетки культивируют при 37°С.
3. При необходимости через 48 ч после заражения культуральную среду заменяют средой, содержащей радиоактивную метку. Например, средой с 2% ЭТС и 1 мкКи/мл – метионина или же средой с 2% ЭТС и 2 мкКи/мл – аминокислот.
4. Первые признаки ЦПД проявляются через 3 дня после инфекции. Затем ЦПД становится более явным, и на 7–10 день после инфекции клетки открепляются от субстрата. Вирус следует собирать именно на этой стадии. Для получения нуклеокапсидов клетки собирают несколько раньше.
Обычно для иммунологических исследований или иммунизации животных мы выделяем CMV из культуральной среды, как описано выше для ВПГ. Однако для анализа спектров вирусных белков и т.д. специалисты рекомендуют использовать метод доктора В. Гибсона, который приводится ниже. Исходным материалом для получения вирусных препаратов максимальной инфекционности и наибольшей чистоты должна быть культуральная среда. Для сохранения высокой инфекционности препарата и целостности вирусных частиц следует избегать осаждения вируса. Впервые центрифугирование в отрицательном градиенте вязкости и положительном градиенте плотности, предложенное Барзилаи и др. для выделения CMV, было применено Тэлботом и Алмейдиа. Метод надежен и гарантирует высокую эффективность. Его осуществляют следующим образом:
1. Клетки собирают в культуральную среду, центрифугируют 10 мин при 1500 g и 4°С, затем наслаивают 3 мл супернатанта на 9 мл градиента 30% глицерина – 35% тартрата. Градиенты готовят согласно.
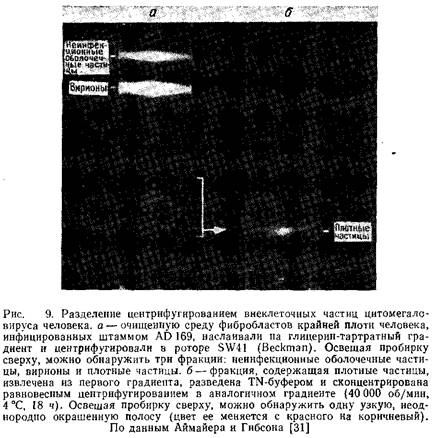
2. Градиент центрифугируют 20 мин при 40000 об/мин и 4°С в роторе SW41. После центрифугирования по светорассеянию, как правило, обнаруживают три фракции: неинфекционные оболочечные частицы, вирионы и плотные частицы. Вирусные частицы каждой фракции могут быть суспендированы в буфере и при необходимости повторно очищены в таком же градиенте.
Гибсон предлагает ряд модификаций этого метода, которые успешно применяются в его лаборатории.
1. Градиенты общим объемом 18 мл готовят в 38-мл пробирках для ротора SW27. На градиент наслаивают 20 мл осветленной культуральной среды и центрифугируют 40 мин при 25000 об/мин и 4°С в роторе SW27.
2. При необходимости обычно используемый буфер трис-НС1 заменяют на 0,04 М фосфатный буфер, рН 7,4.
3. Заменяют тартрат калия на тартрат натрия, если соль калия мешает последующему анализу. Натриевая соль менее растворима при 4°С, чем калиевая.
4. При необходимости вместо тартрата можно использовать градиенты сахарозы. Условия центрифугирования те же.
2.6 Типы частиц CMV
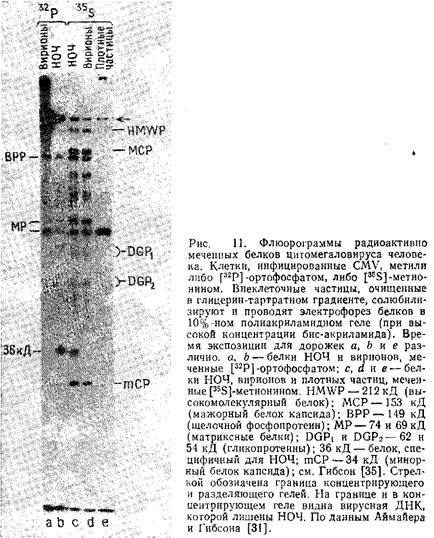
На рис. 10 представлены электронные микрофотографии частиц CMV различной морфологии, полученные методом негативного контрастирования. Спектры структурных полипептидов этих частиц, полученные электрофорезом в полиакриламидном геле, показаны на рис. 11.
2.6.1 Неинфекционные оболочечные частицы
Аймайер и Гибсон провели детальный анализ характерных признаков данных частиц. Несмотря на их сходство по внешнему виду и белковому составу со стандартными вирионами, они не содержат ДНК и, следовательно, неинфекционны.

2.6.2 Частицы высокой плотности
Структура и состав подобных частиц проще, чем неинфекционных оболочечных и стандартных вирионов. Эти частицы представляют собой большие плотные сферы, заполненные гомогенным материалом и окруженные внешней мембраной. Они не содержат ДНК., и 90% всего их белка приходится на матриксный белок, имеющий мол. массу 69 кД.
2.6.3 Вирионы
Гибсон опубликовал подробные спектры структурных белков многих штаммов CMV человека и сравнил их с CMV обезьян. Эти данные представлены на рис. 12 и в табл. 3.
3. Герпесвирус саймири
Герпесвирус саймири можно выделить из крови и культуры клеток большинства здоровых обезьян саймири. Для своих природных хозяев этот вирус не патогенен. Однако он стал предметом пристального внимания ученых после того, как была продемонстрирована его высокая онкогенность для тканей других приматов, в особенности мармозеток из рода Saguinus. У последних в течение 2 мес после заражения развиваются злокачественные опухоли лимфатической системы. Показано, что герпесвирус саймири относится к f-субгруппе герпесвирусов.
3.1 Получение заготовок вируса
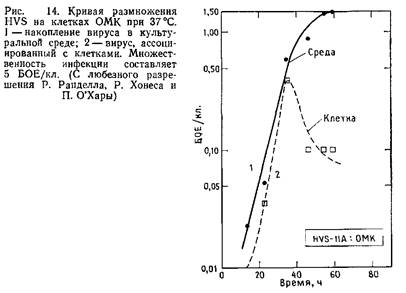
Из большого количества проанализированных клеточных культур для литической инфекции и роста различных штаммов HVS наиболее пригодной оказалась линия клеток почки обезьян дурукули. Кроме того, за небольшим исключением, для получения заготовок HVS можно использовать и клетки линии Vero. Клетки ОМК можно выращивать на большинстве культуральных сред с добавлением ТС.
Для получения заготовок вируса клетки ОМК, растущие в роллерных флаконах, заражают вирусом и культивируют 4–5 дней при 37°С. Оптимальная температура культивирования – 34°С. Однако культивирование при этой температуре может привести к возникновению температурно-чувствительных мутантов. ЦПД проявляется в образовании шарообразных агрегатов клеток, не являющихся истинными синцитиями.
Как правило, ЦПД удается обнаружить на 4–5 день после заражения, хотя эти сроки могут варьировать для различных штаммов. Дочерние вирусы высвобождаются в культуральную среду, при этом в 50 мл среды одного роллерного флакона содержится примерно 2–106 –2-107 БОЕ/мл. Вирусную суспензию с таким титром следует хранить при –70°С, кроме того, вирус можно предварительно сконцентрировать осаждением. Осажденный вирус обычно характеризуют отношением числа физических частиц к инфекционным. Для штамма HVS 11 данное отношение составляет 200–500, этот штамм на редкость стабилен при 4°С. Он теряет за месяц при хранении в культуральной среде всего лишь 0,51g инфекционности.

Таблица 3. Характерные особенности белков CMV
| Сходные белки различных штаммов CMV | Природа белков | Характеристики | |||
| Colburn | CSG | RCMV | HCMV 751 | ||
| 205 | 205 | 207 | 212 | Высокомолекулярный белок | Самый большой белок вириона |
| 163 | 263 | - | 145 | Кислый гликопротеин | Кислый гликозилированный белок |
| 145 | 143 | 150 | 153 | Основной белок капсида | Главный, структурный белок капсида |
| 129 | 129 | 129 | 140 | Белок отсутствует | |
| 119 | 114 | 100 | 149 | Щелочной фосфопротеин | Мажорный фосфорили-рованный, щелочной белок |
| 119 | н. о. | н. о. | – | GP119 | Гликозилированный, кислый |
| 112 | 112 | 112 | 115 | Белок вириона | |
| 100 | н. о. | н. о. | – | GP100 | Гликозилированный, кислый |
| 94 | 90 | 92 | 79 | Предранний | Отсутствует в вирионе, кислый, фосфорилирован |
| 78 | 82 | 75 | 80 | Содержится в вирионе | |
| 69 66 | 69 66 | 69 66 | 74 69 | Белок внешнего матрикса Белок внутреннего матрикса | Мажорный белок, фосфорилирован Мажорный, фосфорилирован, локализуется |
| 65 61 | 64 | 65 | 62 54 57 | Гликопротеин | Мажорный, гликозилированный |
| н. о. | н. о. | Гликопротеин | Гликозилированный | ||
| 59 | 59 | – | GP59, 57 | Гликозилированный | |
| 51 | 50 | 51 | 52 | ДНК-связывающий белок | Фосфорилированный, мажорный, ядерный |
| 40 | 41 | 39 | 42 | Белок вириоиа | |
| 38 | 37 | 39 | 39 | ||
| 37 | 37 | 38 | 35 | Белок сборки | Мажорный, фосфорилированный, локализуется в В-капсидах |

3.2 Титрование вируса
HVS в соответствующих разведениях обычно титруют на монослое клеток Vero или ОМК, растущих в 25 см2 пластиковых флаконах с плоским дном. Вирус титруют методом бляшек. Необходимо отметить, что некоторые партии телячьей сыворотки ингибируют формирование бляшек в культурах ОМК. Во избежание этого сыворотку выдерживают 30 мин при 56°С.
1. Адсорбируют аликвоты соответствующих разведений вируса объемом 1 мл 1–2 ч при 37°С.
2. Аликвоты заменяют на культуральную среду, содержащую 2% ТС и 0,5% КМЦ.
3. Через 7–10 дней инкубации при 37°С начинают формироваться мелкие бляшки. Через 14 дней диаметр бляшек достигает 2–3 мм, хотя, конечно же, данный признак специфичен для каждого штамма.
Немного раньше бляшки формируются в монослойных первичных культурах мармозеток. На рис. 13 показана морфология бляшек HVS через 6 и 9 дней после начала инфекции.
3.3 Цикл размножения
Обычно для получения кривых роста в культуры вносят вирус с высокой множественностью инфекции, а затем через короткие промежутки времени собирают вирус. Таким образом, получают кривую роста HVS. Однако интерпретировать полученные результаты следует с осторожностью. Рэн-делл и Хонес показали, что в этих условиях во многих клетках задерживается инициация вирусной репликации. Если это действительно так, то цикл размножения HVS в одной инфицированной клетке может завершиться за 18 ч.
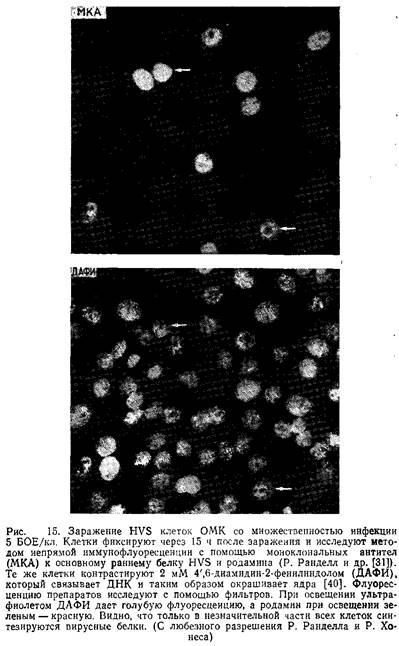
Возможно, что репликация вируса зависит и от клеточного цикла. Эти сложные исследования велись с помощью иммунофлуоресценции на основе моноклинальных антител к антигенам зараженных клеток.
3.4 Получение очищенного вируса
1. Монослойные культуры, выращиваемые в роллерных флаконах, инфицируют с низкой множественностью инфекции, как описано выше. Через 4–5 дней вирус осаждают из культуральной среды центрифугированием при 25000 об/мин 30 мин в роторе SW27. При необходимости через 2 дня после заражения вносят радиоактивную метку.
2. Осадок ресуспендируют в соответствующем буфере. Полученную суспензию по возможности следует оставить на ночь при 4°С.
3. Суспендированный осадок наслаивают на градиент глицерина или сахарозы и центрифугируют при 19000 об/мин 20–30 мин в роторе SW27. После центрифугирования при просмотре пробирок в рассеянном свете обнаруживается одна полоса в центре градиента.
4. Фракцию, состоящую более чем на 90% из вирусных частиц, имеющих оболочку, отбирают, центрифугируют 1 ч при 25000 об/мин и ресуспендируют в соответствующем буфере.
В результате первого градиентного центрифугирования получают осадок, содержащий большое количество вируса. Осадок можно ресуспендировать и дополнительно очистить вторым градиентным центрифугированием. Фракции очищенного вируса могут быть использованы для определения концентрации белка, количества физических частиц и инфекционного титра. Для исследования вирусных частиц электрофорезом в полиакриламидном геле вирус разрушают, как описано выше для ВПГ. Спектр структурных полипептидов HVS штамма 11 показан на рис. 16.