Классификация живых систем
СОДЕРЖАНИЕ: Аспекты разнообразия живых систем. Открытые, закрытые, организменные и надорганизменные живые системы. Первые древнейшие доклеточные протобионты. Адаптивный смысл структурной агрегации монобионтов. Развитие живых систем как функция структурной агрегации.Аспекты разнообразия живых систем
Среди множества разнородных особенностей живых систем наиболее фундаментальные различия между ними создают четыре группы особенностей: 1) общий характер связей живой системы с внешней средой,2) уровень функциональной организации системы, 3) уровень ее структурной агрегации и 4) способ организации процессов метаболизма. Названные аспекты и доставляют естественную основу классифицирования живых систем.
С позиций общей теории систем наиболее принципиальным представляется деление живых систем по характеру их связей с внешней средой, в соответствии с чем выделяются: 1) открытые живые системы, имеющие обмен веществом с внешней средой, и 2) закрытые живые системы, не имеющие такого обмена.
По уровню функциональной организации живые системы естественно подразделяются на 1) организменные живые системы, как элементарные и неделимые, т.е. отдельные организмы, и 2) надорганизменные живые системы, т.е. различные функционально-структурные, ассоциации организмов — колонии, популяции, виды, экосистемы и т. п.
1)По уровню структурной агрегации живые системы делятся на три категории: первичные доклеточные организмы — протобионты, б) простые одноклеточные — прокариоты и одноядерные эукариоты и в) вирусы;
2)метабионтные живые системы, представленные возникшими из мо-нобионтов организмами-метабионтами, у которых генетическая система не централизована в масштабах организма. Таковы многоядерные одноклеточные, ценоцитные и многоклеточные организмы, исключая из числа последних ценометабионтные системы;
3)ценометабионтные живые системы, представленные возникшими из метабионтов организмами-ценометабионтами, организация которых явилась результатом агрегации метабионтных организменных структур. Таковы системы, традиционно называемые «колониями»: сифонофоры, коралловые полипы, мшанки и т. п.
По способу организации процессов метаболизма живые системы делятся на две группы:
1) автобионтные живые системы, представленные организмами, которые метаболируют самостоятельно, располагая необходимыми для этого собственными энергозапасающими, ферментными и синтезирующими аппаратами; к их числу относятся все клеточные организмы;
2) анавтобионтные живые системы, представленные организмами, которые метаболируют только на основе обязательного использования энергозапасающих, ферментных и синтезирующих аппаратов другого организма {живой клетки); таковы все вирусы.
По своей сути рассмотренные четыре деления не адекватны и не соподчинены друг другу, поскольку каждое из них учитывает в развитии живых систем только какую-то одну сторону. Множественность аспектов классифицирования живых систем отражает многогранность процесса их исторического развития.
Открытые и закрытые живые системы
Общие замечания
Традиционное представление о живой системе как имеющей обмен веществом и энергией с внешней средой является недостаточным и неполным, поскольку не охватывает всей целостной картины жизни. Оно соответствует лишь метаболически активным состояниям живых систем и оставляет за рамками живого их метаболически неактивные состояния, определяемые понятием полного, анабиоза, свойственные множеству организмов разных уровней сложности из самых различных систематических групп, от вирусов до высших растений и животных.
Согласно результатам современных исследований, в состоянии полного анабиоза «в организме совсем прекращаются биохимические процессы... происходит приостановка жизненных функций. При этом жизнеспособные структуры находятся в нефункционирующем состоянии, но могут снова возобновить деятельность в благоприятных условиях». В этой связи важно подчеркнуть, что происходящее в состоянии полного анабиоза прекращение метаболических процессов не означает прекращения жизни и, следовательно, не ведет к чередованию в онтогенезе «живых» и «неживых» состояний. Как правильно заметил А. М. Голдовский, «широкое понятие о жизни включает как самый процесс жизни — жизнедеятельность, так и жизнеспособные структуры, функционирование которых составляет этот процесс. Таким образом, понятие о жизнедеятельности является более ограниченным, чем понятие о жизни: жизнедеятельность — это процесс функционирования структур организма, в основе которого лежит сочетание ассимиляции и диссимиляции, имеющий саморегуляторный характер и поддерживающий целостность этих структур, причем структуры в определенные периоды претерпевают изменения и самовоспроизводятся. При анабиозе происходит прекращение жизнедеятельности, а не жизни, поскольку сохраняются жизнеспособные структуры.
В случае полного анабиоза сохраняется только пассивный, т. е. неконтролируемый живой системой обмен со средой, происходящий на тех же основаниях, что и в случае любых тел неорганической природы: происходит, как минимум, принудительный пассивный теплообмен, а в воздушной среде — также и водный обмен, обусловленный определенной гигроскопичностью живых субстратов и естественными изменениями влажности воздуха.
Если находящийся в полном анабиозе организм сохраняет только энергетический обмен со средой в форме пассивного теплообмена, то с позиций общей теории систем его состояние отвечает состоянию закрытой системы. Если же при этом сохраняется также и материальный обмен, в форме пассивного водного обмена или в какой-либо иной пассивной форме, то состояние организма, как и в случае его выраженной метаболической активности, формально должно было бы быть определено как состояние открытой системы. Принимая, однако, во внимание, что этот пассивный материальный обмен, весьма односторонний по содержанию и незначительный по объему, в данном случае осуществляется не организмом как целостной живой системой, а лишь субстратами, из которых он состоит, следует признать, что и в этом варианте полного анабиоза состояние организма фактически отвечает таковому закрытой системы.
Следовательно, обменные процессы, осуществляемые метаболически активным организмом, и процессы пассивного обмена, которые испытывает метаболически инертный организм, находящийся в состоянии полного анабиоза, принципиально различны. Первые отличаются глубокой качественной специфичностью и системной комплексностью, причем их содержание и объем всецело определяются конкретными метаболическими потребностями организма и постоянно находятся под его контролем. Вторые, напротив, спонтанны, разрозненны и не контролируются организмом, будучи обусловлены лишь соотношением физико-химических свойств биологических субстратов и окружающей их среды. Эти различия и определяют основания к тому, чтобы в каждом конкретном случае трактовать организм как открытую или закрытую систему: если метаболически активный организм принципиально возможен только как открытая система, то метаболически инертный, т. е. находящийся в состоянии полного анабиоза, биологически представляет собою закрытую систему, поскольку не имеет закономерного материального обмена со средой.
Таким образом, привычные «представления об организме как открытой системе не могут охватывать все его состояния, и в том числе анабиоз. Ведь при переходе к полному анабиозу вследствие приостановки жизнедеятельности порываются обычные связи со средой, организм)Хклет-ка) перестает быть открытой системой в обычном понимании, поскольку прекращается обмен веществ. Разумеется, даже при анабиозе может идти обмен веществом и энергией с окружающей средой за счет поглощения или испарения воды, повышения или; понижения температуры, но такие изменения характерны также и для куска любого набухающего, гидрофильного геля при колебании влажности и температуры среды».
В общей теории систем наряду с «закрытыми» различают системы «изолированные»; различия между ними заключаются в том, что первые обмениваются со средой энергией, но не имеют обмена веществом, тогда как вторым свойственно отсутствие обмена как веществом, так и энергией. Совершенно ясно, что понятие изолированных систем если и прилржимо к живым системам, то только в сравнительно редких случаях, поскольку полное отсутствие пассивного энергетического обмена со средой возможно только в условиях полной изотермии. Таким образом, живые системы могут находиться как в состоянии открытых, так и в состоянии закрытых и изолированных систем. Учитывая, однако, малую вероятность изолированного состояния живой системы, целесообразно выделять два заведомо существующих состояния: открытые и закрытые живые системы.
Различия, открытых и закрытых живых систем имеют принципиальное значение при рассмотрении особенностей экоморф и находят отражение в структуре единой экоморфологической системы организмов, что детальнее рассматривается в гл. 7.
Открытые живые системы
Открытая живая система метаболически активна и существует на основе постоянного двустороннего материального и энергетического обмена с внешней средой, причем со стороны живой системы этот обмен является активным в том смысле, что его содержание и объем структурно обусловлены системой на молекулярном и надмолекулярном уровнях, т. е. находятся под контролем системы и соответствуют ее метаболическим потребностям, будучи, в частности, определены необходимостью поддержания положительного энергобаланса системы.
Этому активному обмену живых открытых систем должен бы ь противопоставлен свойственный неживым открытым системам пассивный обмен веществом и энергией, который определяется только совокупностью физико-химических свойств системы и окружающей ее среды. К числу таких пассивных обменных процессов относятся, например, теплообмен между твердым телом и окружающей его подвижной жидкой средой, химическое взаимодействие поверхности этого тела с обтекающей его жидкостью и т. п.
Если живые открытые системы в результате двустороннего материального и энергетического обмена со средой сохраняют свою квазистационарность, то системы неживые под действием этого обмена закономерно разрушаются, поскольку их изменения подчиняются лишь второму закону термодинамики и ведут к общему уменьшению свободной энергии всей макросистемы. В этом — принципиальное различие живых и неживых открытых систем.
Метаболическая функция свойственна открытым живым системам всех уровней сложности и отражает общее для них свойство саморазвития. С позиций общей теории систем открытая живая система может быть определена как такая, которая в ходе взаимодействия с окружающей ее средой «сохраняется неизменной при постоянном изменении ее составных частей».
На организменном уровне имеет место постоянное самообновление открытой живой системы на основе свойственного всем организмам обмена веществом и энергией с внешней средой. На уровне популяций и видов эта метаболическая функция обеспечивается как индивидуальным метаболизмом отдельных организмов, так и сменой организмов в популяции в ходе естественной смены поколений. На уровне экосистем и биогеоценозов в филогенетическом плане ко всему этому присоединяются еще смена видов и изменение их соотношений, происходящие на основе естественного развития конкретных сообществ под действием сложной совокупности разнообразных факторов.
При всем видимом разнообразии конкретного содержания и форм саморазвития живых систем различного уровня сложности сущность этого процесса во всех случаях принципиально одна и та же и заключается именно в сохранении известной квазистационарности системы при постоянном фактическом изменении ее составных частей, что в конечном итоге всегда представляет собою результат реализации конкретных -филогенетически обусловленных программ развития, имеющих материальное выражение в геномах конкретных организмов. Иначе говоря, развитие живых систем любого уровня сложности всегда определяется генетической информацией, которая содержится в материализованной форме на организменном уровне.
Таким образом, открытые живые системы всегда представлены организмами, находящимися в метаболически активном состоянии. Филогенетически это состояние, несомненно, является исходным для первичных организмов, поскольку они могли возникнуть не иначе как метаболически активное начало, противостоящее энтропии.
Закрытые живые системы
Закрытая живая система метаболически неактивна, ей свойственно полное отсутствие активно осуществляемого материального обмена с внешней средой. Это определяет отсутствие всех прочих жизненных функций и сводит жизненный процесс к существованию нефункционирующей структурной системы организма, что соответствует состоянию полного анабиоза. Таковы, например, вирионы вирусов, эндоспоры бацилл, семена многих растений, высохшие коловратки и тихоходки в состоянии ксероанабиоза и т. п. Реальную основу существования подобных закрытых живых систем составляют их эволю-ционно возникшие структурные антиэнтропические свойства, позволяющие структурной системе организма сохранять свою жизнеспособность в метаболически неактивном состоянии, при отсутствии положительного баланса в энергообмене со средой.
В ряде случаев организм может находиться в состоянии закрытой системы весьма продолжительное время, намного превосходящее обычную длительность активного онтогенеза. Достаточно сказать, что после сотен и даже тысяч лет пребывания в анабиотическом состоянии обнаруживают нормальную жизнеспособность споры некоторых бактерий, и семена многих растений. Столь длительное сохранение жизнеспособности нефункционирующей структурной системы организма возможно только потому, что полный анабиоз представляет собою не «замедленную жизнедеятельность» с вытекающей отсюда необходимостью каких-то пусть минимальных, но неизбежных энергозатрат, как думали прежние исследователи, а полное отсутствие жизнедеятельности и, соответственно, полное отсутствие энергозатрат.
Антиэнтропические свойства структурной системы организма, проявляющиеся в состоянии полного анабиоза, эволюционно обусловлены на молекулярном и надмолекулярном уровнях и, несомненно, представляют собою один из важнейших комплексов адаптации, обеспечивающих сохранение живой системы в меняющихся условиях среды. В разных царствах органического мира существует множество вариантов подобных адаптации, из которых мы остановимся лишь на двух примерах: 1) вирионах вирусов и 2) эндоспорах бацилл и некоторых актиномицетов.
Вирионы — метаболически инертная внеклеточная стадия в онтогенезе вирусов. Не имея собственного обмена веществ и не проявляя никаких других признаков жизни, они способны испытывать лишь деструктивный метаболизм, разрушаясь под действием различных неблагоприятных факторов как внутри организма хозяина, так и вне его. Как одна из стадий онтогенеза облигатных внутриклеточных паразитов, каковыми являются все вирусы, вирионы чаще всего существуют либо в средах биологического происхождения, либо в средах, оптимальных для развития организма-хозяина, т. е. в таких средах, параметры которых далеки от экстремальных для жизни. Поэтому, в отличие от большинства случаев анабиоза клеточных организмов, состояние полного анабиоза вирионов, в котором они находятся вне живой клетки, не может быть объяснено высокой агрессивностью среды.
По мнению автора, причина анабиотического состояния вирионов заключается в «минимальности» общей конструкции организма вируса и глубоком своеобразии свойственного вирусам способа организации процессов метаболизма. Существенной чертой вирусов является их метаболическая несамостоятельность — полная зависимость активной фазы их онтогенеза от деятельности энергозапасающих, ферментных и синтезирующих аппаратов живой клетки.. На этом основании в единой экоморфологической системе организмов вирусы выделены в особое царство «анавтобион», которое противостоит царству «автобион», объединяющему экоморфы клеточных организмов. Именно метаболическая несамостоятельность вирусов, эволюционно возникшая как результат глубокой всесторонней функционально-структурной интеграции их с живой клэткой, и обусловливает полную метаболическую инертность вирионов: как минимальные живые структуры, они экологически представляют собою лишь способ сохранения во внеклеточной среде и доставки к новому хозяину вирусных геномов, необходимых для обеспечения синтеза новых поколений вируса.
Антиэнтропические свойства вирионов соответствуют особенностям среды, в которой они существуют, и прежде всего, тому факту, что большинство внутренних сред организма, обычно служащих ареной развития вирусов, содержат в тех или иных концентрациях нуклеазы и протеазы, оказывающие литическое действие на нуклеиново-белковые субстраты вирионов. Важнейшим.структурным антиэнтропическим фактором вирио-на является его белковый капсид, основное назначение которого состоит в предохранении содержащейся в вирионе нуклеиновой кислоты от инактивации ее присутствующими в среде нуклеазами.
Устойчивость вирусного капсида к действию протеолитических ферментов в значительной мере определяется его макроструктурой и только в меньшей степени зависит от первичной структуры капсидного белка. Сама по себе пептидная цепь вирусного белка в изолированном виде не имеет каких-либо особенностей в своей первичной, вторичной и третичной структуре и поэтому легко гидролизуется протеолитическими ферментами. Устойчивость к этим ферментам обусловлена макромолекулярной структурой белковых субъединиц капсида, при которой пептидные связи, непосредственно испытывающие на себе действие ферментов, оказываются для них недоступными. Примером может служить построенный по принципу спиральной симметрии вирион вируса табачной мозаики, в составе которого, благодаря специфической укладке пептидных цепей, их концы, откуда начинается гидролиз белка экзопротеазами, находятся, как правило, в глубине капсида. При нарушении целостности капсида вирион сразу же приобретает чувствительность к клеточным протеазам, что становится целесообразным при попадании его в клетку, где вирусный геном должен быть освобожден от капсида.
Эндоспоры бацилл и некоторых актиномицетов, как и вирионы вирусов, метаболически полностью инертны. Однако если в случае вирионов эта инертность обусловлена свойственной всем вирусам метаболической несамостоятельностью и сама по себе не связана с особенностями внешней среды, параметры которой, как правило, вполне пригодны для жизни, то в случае эндоспор метаболическая инертность представляет собою адаптацию, направленную именно на предохранение организма от губительного действия резко агрессивных факторов среды. И в этом смысле именно эндоспоры бацилл и актиномицетов служат непревзойденным образцом антиэнтропической стойкости, сохраняя свою жизнеспособность: при воздействии таких остро агрессивных факторов, как глубокое обезвоживание, колебания температуры в диапазоне от —250 до +170 °С, ионизирующие излучения, вакуум, действие концентрированных кислот и ферментов и др..
Аналогия в химической и макроструктурной специфике эндоспор микроорганизмов двух различных групп — бацилл и некоторых актиномицетов —. сама по себе представляет значительный интерес. Она проявляется, в частности, в том, что в обоих случаях эндоспоры имеют следующие общие особенности: 1) накапливают дипиколиновую кислоту; 2) имеют близкое суммарное содержание гуанина и цитозина в ДНК, причем в этом отношении Thermoactinomyces и Actinobifida существенно отличаются от других э у актиномицетов, у которых ГЦ-со-держание значительно выше; 3) образуют сходные структуры в оболочке, обнаруживая при этом аналогичную устойчивость к нагреванию и высушиванию. В этом следует, возможно, видеть не генетическую близость Thermoactinomyces и Actinobifida к Bacillaceae, а проявление особенностей определенной экоморфы, т.е. эндоспоры, конвергентно возникшей у бацилл и актиномицетов. Такой взгляд на рассмотренную аналогию эндоспор этих организмов кажется тем более допустимым в свете общеизвестных существенных морфологических различий их вегетативных форм.
Антиэнтропические свойства эндоспоры структурно обусловлены как ее химическим составом, отличным от такового вегетативной клетки, так и макроморфологией ее оболочки, т.е. определены на молекулярном и надмолекулярном уровнях.
На макроморфологическом уровне жизнестойкость метаболически неактивной эндоспоры обусловлена наличием сложно устроенной мощной оболочки, содержащей такие структурные элементы, которые отсутствуют у вегетативной клетки. Внутренняя, прилежащая к протопласту зона оболочки образована двумя слоями обычной клеточной мембраны, между которыми развивается специфичная для эндоспоры толстая прочная электронопрозрачная оболочка — кортекс, состоящая из пептидогликана уникальной структуры, возможно, сходной для всех или большинства спорообразующих бактерий и во многих отношениях отличной от структуры пептидогликанов вегетативных клеток. Внешняя зона оболочки в основном белковая, содержащая до 80% всех белков споры, причем эти белки отличаются необычно высоким содержанием цистеина и гидрофобных аминокислот и обнаруживают чрезвычайную устойчивость к различным литическим факторам.
Дипиколиновая кислота, содержащаяся в эндоспоре в виде дипико-лината кальция, составляет 10—15% сухой массы споры и локализована в протопласте; она обусловливает термостабильность зрелой споры, что показано экспериментально. Другой важный химический фактор жизнестойкости эндоспор — их сильная обезвоженность; с увеличением содержания воды в спорах терморезистентность их снижается.
Таким образом, примеры вирионов и эндоспор показывают, что возможность существования закрытых живых систем всецело обусловлена структурными адаптациями организмов на молекулярном и надмолекулярном уровнях. Эти адаптации позволяют нефункционирующей, метаболически инертной структурной системе организма в течение некоторого времени сохранять свою жизнеспособность, уклоняясь от активного, функционального контакта с окружающей средой, параметры которой в той или иной мере выходят за пределы эволюционно обусловленного диапазона, пригодного для метаболически активной жизни.
Следовательно, закрытые живые системы всегда представлены организмами, находящимися. в метаболически неактивном состоянии, т. е. в состоянии полного анабиоза.
Организменные и надорганизменные живые системы
Деление живых систем на организменные и надорганизменные отражает два основных типа их функциональной организации. Это деление по своей сути не имеет адекватных ему альтернатив: как функционально неделимая, генетически первичная живая система, имеющая собственную материализованную программу развития в форме генома, организм принципиально отличается от любых надорганизменных ассоциаций, программа развития которых всегда обеспечена только через посредство их подсистем, т. е. конкретных организмов, образующих ассоциацию. В силу этого, по сравнению с делением живых систем на организменные и надорганизменные, все подразделения последних могут иметь только подчиненное значение.
Поэтому, полностью разделяя мнение о том, что в иерархии уровней организации живого организменный уровень является основным, универсальным и первичным, автор считает, что при делении всего многообразия существующих живых систем по уровням их организации как первую ступень классификации следует выделять организменные и надорганизменные системы. Выделение же в качестве равнозначных по рангу трех основных уровней — орга-низменного, популяционного и экосистемного в свете сказанного представляется необоснованным.
Организменный и надорганизменный уровни функциональной организации живой системы наиболее существенно различаются 1) по информационной структуре живых систем, 2) по степени их делим сти и 3) по степени иерархичности их функционально-структурно организации.
Информационная структура организменных и надорганизменных живых систем принципиально различна. Все организмы характеризуются наличием централизованной собственной программы развития, основу которой составляет геном, содержащий определенный объем конкретной генетической информации. Централизация собственной программы развития определяет функционально-структурную целостность организма как неделимой единицы жизни. В надорганизменных системах программа развития не является централизованной и существует лишь как интегральный генофонд, образованный геномами конкретных организмов.
Эти различия организменных и надорганизменных систем в структуре собственной программы развития имеют фундаментальное значение в определении всех их прочих особенностей, и прежде всего степени их функционально-структурной интеграции, общий уровень которой в организменных системах всегда неизмеримо выше, чем в надорганизменных. В этом отношении между организменными и надорганизменными системами существуют качественные различия, определяемые мерой их делимости.
Делимость организменных и надорганизменных живых систем различна в принципе. Организмы всегда принципиально неделимы, что с неизбежностью вытекает уже из самого определения организма как элементарной живой системы, тогда как надорганизменные системы — популяции, колонии, экосистемы и т. п.,— напротив, в той или иной мере всегда делимы.
Если неделимость организма-монобионта в силу моноцентричности его генетической структуры всегда представляется достаточно очевидной и не требует каких-либо комментариев, то неделимость метабионтов и це-нометабионтов в свете известных фактов их вегетативного размножения на первый взгляд часто выглядит спорной. Достаточно вспомнить хотя бы, что,из фрагмента листа бегонии вырастает целое растение, растертая в ступке гидра превращается во множество новых гидр, а из тополя, разделив его на черенки, можно вырастить целую тополиную рощу.
В действительности, однако, подобные факты не противоречат идее неделимости организма и в то же время не свидетельствуют о том, что потенциально способный к вегетативному размножению родительский организм является составным. Вся суть дела заключается в том, что отделенные от родительского организма фрагменты сразу же начинают контактировать со средой как целостные организменные живые системы, что было не свойственно им до отделения от родительской живой системы, в пределах которой они проявляли себя лишь как безусловно несамостоятельные, в функциональном отношении односторонне специализированные структуры. В этом проявляется пластичность информационной структуры многоклеточных организмов, обусловленная тем, что каждая клетка многоклеточного тела имеет собственный геном, благодаря чему потенциально является зачатком целостной организменной системы. Поэтому механическая делимость тела, характерная для некоторых многоклеточных, не- означает функциональной делимости их как организменных систем: при насильственном, эктогенном делении тела такого организма возникающие фрагменты именно вследствие отделения их от родительской системы сразу же превращаются в дочерние организмы, т. е. вступает в действие эволюционно выработанный механизм вегетативного размножения с помощью простой фрагментации.
При этом, однако, для каждого многоклеточного организма, способного к вегетативному размножению посредством эктогенной фрагментации, существует определенный минимальный размер фрагмента, при котором он еще может взаимодействовать со средой как целостная организменная система. Например, если уже упомянутая бегония может размножаться даже фрагментами листьев, то для тополя минимальным жизнеспособным фрагментом является стеблевый черенок, т. е. кусочек стебля с небольшим числом листьев. Это указывает на эволюционную обусловленность допустимых уровней эктогенного фрагментирования и подтверждает правильность его интерпретации как способа вегетативного размножения: слишком мелкие фрагменты, размер которых ниже допустимого для данного вида, оказываются уже нежизнеспособными и здесь мы можем, следовательно, убедиться в том, что и в подобных случаях сохраняется обычная для всех организмов функциональная неделимость.
Таким образом, в любом случае вегетативного размножения мы можем констатировать принципиальную неделимость организма за пределами того эволюционно обусловленного уровня фрагментирования, который является экологически целесообразным для данного вида.
Иерархичность организации в той или иной мере свойственна всем живым системам. Однако если на организменном уровне она всегда хорошо выражена и составляет главное условие, обеспечивающее возможность структурного усложнения организмов, то в надорганиз-менных системах в некоторых случаях может быть выражена, напротив, очень слабо, примером чего могут служить хотя бы разного рода временные ассоциации животных, возникающие на относительно короткое время, в частности такие, как небольшие стайки мелких кочующих птиц.
Организменные живые системы
Организменные живые системы представляют собою филогенетически первичный, элементарный вариант живой системы, возникновение которого исторически соответствовало началу собственно биологической эволюции и само по себе явилось результатом длительного предбиологичес-кого развития.
Как автокаталитический процесс предбиологическая эволюция, судя по всему, имела в своей основе принцип гиперцикла, т. е. «принцип естественной самоорганизации, обусловливающий интеграцию и согласованную эволюцию системы функционально связанных самореплицирующихся единиц». Такими самореплицирующимися единицами на ранних стадиях предбиологической эволюции были, видимо, предшественники рибонуклеиновой кислоты — РНК-подобные полимеры, которые «в силу своих физических свойств наследуют способность к самовоспроизведению, а это является необходимой предпосылкой для систематической эволюции».
С возникновением процесса трансляции в этот автокаталитический гиперцикл были вовлечены аминокислоты, в результате чего он стал нук-леиново-пептидным, а в конечном счете, после достаточного удлинения пептидных цепей,— нуклеиново-белковым, соединив в себе, таким образом, оба важнейших химических компонента будущей организменной системы: информационное начало в виде нуклеиновой кислоты и структурно-каталитическое — в виде белка.
Не останавливаясь на деталях предбиологической эволюции, отделенной от нас почти 4 млрд. лет и происходившей в условиях, о которых мы не имеем достаточно ясного представления, отметим лишь, что на этом предбиологической, т. е. доорганизменном, этапе эволюционирующий функционально-структурный комплекс был представлен некоторой ассоциацией доорганизменных систем — макромолекул и простых надмолекулярных агрегатов. Интегративная эволюция подобных ассоциаций со
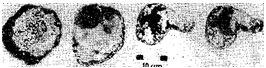 |
Рис. 1 Древнейший известный организм — бактерия Isuasphaeraisua. По Pflug, с изменениями.
временем привела к становлению первых простых организмов — прото-бионтов. Известные в настоящее время наиболее древние реальные палеонтологические свидетельства существования таких организмов обнаружены в отложениях, возраст которых приближается к 4 млрд. лет. Таковы, в частности, мельчайшие сфероподобные организмы диаметром около 30 мкм, найденные в Юго-Западной Гренландии в слоях возрастом около 3,8 млрд. лет и описанные как Isuasphaeraisua.
Таким образом, первичные организмы-протобионты возникли на основе длительной предбиологической интегративной эволюции некоторой ассоциации доорганизменных структур, организованных на уровне макромолекул и простых надмолекулярных агрегатов. Становление протобион-тов явилось непосредственным результатом акта структурной агрегации этих доорганизменных объектов молекулярного уровня, и прежде всего молекул нуклеиновых кислот и белков, воссоединение которых в единую автокаталитическую самореплицирующуюся информационно-структурную систему было, несомненно, самым главным, первым и решающим шагом на пути превращения ассоциации доорганизменных структур в организм.
Как функционально неделимые самореплицирующиеся информационно-структурные системы все первичные организмы имеют одну важнейшую общую черту: они элементарны по своей информационной структуре. Все они имеют только один информационный центр в форме генома, т. е. одну единую, неделимую материализованную генетическую программу развития, всегда централизованную в масштабах организменной живой системы. В соответствии с этим, принимая во внимание информационную неделимость подобных организмов, мы называем ихмонобионтами, а их единую, неделимую собственную программу развития — монобионтной.
Монобионтами были первые древнейшие доклеточные протобионты. Среди современных организмов к числу монобионтов относятся: 1) простые одноклеточные — прокариоты и одноядерные эукариоты и 2) вирусы, представляющие собою, судя по всему, результат вторичного упрощения более сложных, организмов. Если монобионтный характер одноклеточных прокариот и вирусов всегда достаточно очевиден, то мо-нобионтность одноядерных эукариот нуждается в специальном обсуждении в связи с 1) вероятным симбиотическим происхождением эукариоти-ческой клетки, 2) ее различной плоидностью и 3) фрагментацией ее ядра.
Симбиотический характер эукариотической клетки в настоящее время признается почти всеми. Если классическая точка зрения предполагает аутогенное происхождение всех эукариотических органелл, т. е. возникновение их из вещества протоэукариотической клетки, то, согласно симбиотической теории, эукариотическая клетка возникла в результате серии последовательных симбиозов. Суть этой теории сводится к тому, что «всё эукариоты •сформировались в результате симбиоза между чрезвычайно далекими друг от друга видами прокариот: нуклеоцитоплазма образовалась из микроорганизмов- «хозяев», митохондрии — из бактерий, дышащих кислородом, пластиды — из хлорокси- или цианобактерий, а ундулиподии — из спирохет, прикреплявшихся к поверхности хозяев». «Если гетеротрофные эукариоты три-геномны благодаря своим митохондриям, ундулиподиям и нуклеоцитоплаз-ме, то растения уже квадригеномны благодаря дополнительной органел-ле — своим фотосинтезирующим пластидам».
Таким образом, согласно симбиотической теории, в результате серии последовательных симбиозов возникли одноклеточные эукариотические организмы, давшие затем начало всему многообразному миру многоклеточных эукариот. Не рассматривая конкретные аргументы этой теории, которые излагаются в книге Л. Маргелис, заметим, что симбиотиче-ская природа эукариотической клетки представляется особенно вероятной в свете четырех обстоятельств, каковы:
1) многочисленные и весьма разнообразные по характеру факты эндо-симбиоза современных прокариот и одноклеточных эукариот;
2) гомология нуклеотидных последовательностей в пределах отдельных категорий органелл, более выраженная между аналогичными органеллами различных видов и групп эукариот, чем между любой из органелл и «своей» нуклеоцитоплазмой данного вида;
3) глубокие и разноплановые морфологические гомологии в пределах каждой из категорий органелл, примером чего могут служить, в частности, такие факты, как структура эукариотических ундулиподии, построенных по типу 9 + 2, и центриолей и кинетосом — по типу 9 + 0;
4) многочисленные и глубокие аналогии между органеллами современных эукариот и свободноживущими прокариотами, прежде всего аналогии между хлоропластами и различными цианобактериями.
Хотя симбиотическая концепция эукариотической клетки, несомнен-.но, основывается на множестве неопровержимых фактов и серьезных логических доводов, наряду с нею продолжает существовать и противоположная точка зрения, утверждающая идею прямой филиации, т. е. несим-биотический генезис эукариот. Для нас, однако, в связи с рассмотрением особенностей монобионтов наибольший интерес представляет именно симбиотическая теория: идея прямой филиации уже сама по себе находится в явном и бесспорном соответствии с монобионтным характером одноядерной эукариотической клетки, тогда как симбиотическая концепция, утверждающая гетерогенный характер одноядерной одноклеточной эукариоты, требует специального обсуждения.
В действительности же и допущение симбионтной природы одноядерной эукариотической клетки нисколько не противоречит ее монобионтному характеру. Каково бы ни было происхождение эукариотических органелл — митохондрий, пластид и ундулиподии,— их присутствие в. одноядерной клетке не нарушает моноцентричности ее генетической структуры, поскольку это свойство всецело обусловлено наличием в ней единственного ядра. Как бы ни были многочисленны и разнообразны внеядерные геномы, вся их совокупность всегда находится в тесной функциональной связи •с ядерным геномом, который является важнейшим и единственным координирующим генетическим центром, что и определяет монобионтный характер одноядерной клетки и ее собственной программы развития.
Соответственно этому типы клеток — монады, диады, триады, тетрады, пентады, гексады и т. п., различающиеся по числу содержащихся в них белоксинтезирующих единиц, при условии их одно-ядерности все являются монобионтами, поскольку их генетическая структура остается строго централизованной благодаря наличию в каждой из них единственного ядра.
Плоидностъ одноядерной клетки, т. е. наличие в ней одной, двух или же четырех и более копий генома, также не меняет ее общей централизованной информационной структуры, характерной для монобионтов.
Диплоидность создает, как известно, значительные биологические преимущества, позволяя, в частности, нейтрализовать многие вредные мутации, поскольку большинство из них рецессивны и; в диплоидных клетках подавляются доминантными нормальными аллелями. За счет диплоидности достигается, следовательно, стабилизация, генетической программы развития клетки при сохранении строго моноцентрического характера ее информационной структуры. Наличие этого-защитного дублирующего механизма подчеркивает функциональную целостность диплоидного ядра и подтверждает тем самым принципиальное тождество моноцентрической информационной структуры гаплоидных и диплоидных клеток.
Полиплоидность особенно обычна у растений. Она способствует увеличению размеров организма и повышает его устойчивость к разного рода неблагоприятным условиям, что, в частности, проявляется в увеличении доли полиплоидных видов растений в арктических и высокогорных областях. Например, доля полиплоидных видов во всей наземной растительности увеличивается с 37% на Кикладах и 38% в Алжире до 76% на Шпицбергене и 86% в Северной Гренландии. Полиплоидность изменяет, следовательно, некоторые свойства ядра как информационного центра клетки, однако не нарушает при этом моноцентрического характера ее общей информационной структуры: при любом уровне плоидности клетка остается одноядерной и, следовательно, генетически моноцентричной.
Известные различия между гаплоидными, диплоидными и полиплоидными клетками имеются в делимости их ядер, однако эти различия-не касаются генетического моноцентризма клеток. Например, только для-полиплоидной клетки возможен такой саособ деления, при котором ее-высокополиплоидное ядро делится на некоторое число ядер с более низко» плоидностью и образуется соответствующее число дочерних клеток, как. это наблюдается, например, у некоторых радиолярий.
Таким образом, диплоидность и полиплоидность способствуют повышению жизнеспособности организма путем увеличения его размеров и? повышения стабильности его генетической системы. Функциональная и структурная целостность ядра с повышением уровня его плоидности не только не нарушается, но и становится более глубокой и многоплановой, т. е. моноцентризм генетической системы клетки дополняется новыми аспектами. Следовательно, генетический моноцентризм, т. е. функциональная неделимость информационной структуры в равной мере свойственна-всем одноядерным клеткам.— как гаплоидным, так и диплоидным и полиплоидным.
Поэтому все одноядерные одноклеточные, независимо от степени плоидности их ядер, принадлежат к числу монобионтов, т. е. имеют строго-моноцентрическую информационную структуру.
Структурная фрагментированностъ ядра эукариотической клетки также не нарушает ее генетического моноцентризма, если совокупность всех фрагментов образует функционально неделимый комплекс. Классическим примером такой ситуации может служить ядерный дуализм инфузорий. Два ядра инфузорий — макронуклеус и микронуклеус — имеют различные функции и представляют собою элементы функционально единого, неделимого генетического аппарата, лишь морфологически разделенного на два фрагмента, один из которых выполняет функции, связанные с синтезом белка, а другой — генеративные. В связи с этим у инфузорий комплекс из двух ядер-следует, по мнению автора, считать единым морфологически фрагменти-рованным ядром; при наличии одного фрагментированного ядра, т. е. одного макронуклеуса и одного-микронуклеуса, как у Paramecium, генетическая программа инфузории фактически является столь же централизованной, как и у «обычных» одноядерных простейших. Поэтому инфузории также, как и все одноядерные простейшие, относятся к числу монобионтов.
В случае 1 многоядерности одноклеточных эукариот совокупность имеющихся в клетке ядер с функциональной стороны может представлять собою один.из двух возможных вариантов: либо эти ядра образуют функционально неделимый комплекс, т.е. единый морфологически фрагменти-рованный информационный центр, как у Paramecium, либо они относятся друг к другу как идентичные и автономные информационные центры, как у жгутиконосцев из группы Opalinina. В первом случае организм является монобионтом, а во втором — относится к числу метабионтов.
Монобионты — элементарные блоки жизни, простейшие живые системы организменного уровня. Уже на заре биологической эволюции эта элементарная, монобионтная живая система, пройдя первичную, докле-точную фазу своего развития, обрела форму биологической клетки, ставшей основным структурным блоком в последующем агрегатном усложнении живых систем. В иерархическом ряду уровней функциональной организации живых систем монобионтный уровень является, таким образом, самым первым, наиболее низким, и притом первым организменным уровнем организации живого. Ему соответствует и самая простая из возможных — неделимая информационная структура живой системы, материально организованная на субмолекулярном уровне и представленная геномом.
Первый ррганизменный уровень функциональной организации живой системы соответствует ее элементарной организации, которая в функциональном, структурно-энергетическом и информационном отношениях представляет собою минимальный реально возможный вариант. С особенной четкостью это проявляется в моноцентризме информационной структуры системы, что находит конкретное выражение в неделимости ее генетической программы и полной централизации этой программы в масштабах организма-монобионта.
Таким образом, у всех монобионтов — прокариот, одноядерных одноклеточных эукариот и вирусов — программа развития неделима и централизована на генетическом уровне, что принципиально отличает их от всех других организмов.
Адаптивная эволюция клеточных живых систем привела к ряду принципиальных усложнений в их функциональной организации. Централизованный характер программы развития системы при этом сохранился, но принципы и степень этой централизации претерпели существенные изменения.
Если изменения плоидности клетки не нарушают моноцентрического характера ее информационной структуры, то в многоядерной клетке, например у многоядерных одноклеточных жгутиконосцев, генетическая программа уже не является централизованной в масштабах клетки, поскольку каждое из ядер пространственно вполне обособлено. В этом случае централизованный характер собственной программы развития многоядерного одноклеточного организма обусловлен уже не на первичном, т.е. генетическом, уровне, а на уровне межъядерных взаимодействий, которые обеспечивают развитие организма как целостной живой системы. Аналогичное положение характерно для всех цеиоцитных организмов, многоядерные тела которых не разделены на клетки. К их числу относятся, в частности, многоядерные протоплазменные плазмодии слизевиков, примитивные грибы с неоептированным, т. е. ценоцитным, мицелием и сифоновые водоросли. У всех подобных организмов каждое ядро вместе с прилегающей к нему зоной протоплазмы образует энергиду, т. е. в известной мере генетически автономную область, которую можно рассматривать как «сферу влияния» конкретного ядра. Централизация собственной программы развития у таких ценоцитных организмов обеспечивается на основе интеграции генетических программ, содержащихся в отдельных ядрах, т. е. на уровне межъядерных взаимодействий.
Ценоцитная организация имеет много общего с многоядерной одноклеточной: в обоих этих случаях организм представляет собою многоядерный агрегат, не поделенный на клетки. В этом смысле явно одноклеточные многоядерные зоожгутиконосцы из группы Polymastigina и Opalinina принципиально не отличаются от неклеточных многоядерных плазмодиев слизевиков и многоядерных неклеточных мицелиев грибов из групп Chytridiomycetes, Hyphochytriomycetes, Oomycetes и Zygomycetes или же многоядерных неклеточных талломов сифоновых водорослей из группы Caulerpaceae, которые обычно называют ценоцитными. Макро-структурные различия между всеми этими организмами носят в основном количественный характер и определяются прежде всего их линейными размерами и уровнем сложности как многоядерных систем — в первую очередь числом ядер и степенью функциональной и структурной обособленности отдельных одноядерных участков тела, т. е. отдельных энергид. При любом числе ядер, любой степени структурной сложности и любой автономии энергид не поделенные на клетки многоядерные организмы могут иметь принципиально различный генезис, поскольку могут возникать 1) в результате деления ядра в исходно одноядерной клетке, как это происходит, например, у зоожгутиконосцев из групп Opalinina или Polymastigina, либо 2) путем слияния в единый ценоцитный агрегат множества изначально обособленных одноядерных клеток, что можно видеть, в частности, на примере плазмодиев слизевиков из группы Асга-siomycetes.
К числу общих особенностей многоядерных одноклеточных и цено-цитных организмов относится и свойственная им способность к вегетативному размножению путем деления многоядерного тела на.фрагменты, каждый из которых содержит некоторое число родительских ядер.
Ясно, что между многоядерными одноклеточными и ценоцитными организмами невозможно провести резкую границу, В то же время разграничение их целесообразно, поскольку отражает степень их структурной сложности как многоядерных систем. Поэтому в нижеследующем мы будем понимать под много ядерными одноклеточными относительно небольшие организмы, макроформа которых близка к наиболее обычной форме клетки, т. е. характеризуется сравнительной простотой и отсутствием выраженного разветвления; организмы же крупные и разветвленные будем называть ценоцитными Несмотря на явную условность подобного деления, оно, несомненно, способствует конкретизации объекта и тем самым уточняет изложение.
Многоклеточность, по сравнению с ценоцитной многоядерностью, представляет собою дальнейшее усложнение агрегатного состояния живой системы и, соответственно, ведет к дальнейшему усложнению собственной программы ее развития. Каждая клетка многоклеточного организма имеет собственный геном; поскольку геномы во всех клетках идентичны, все клетки организма генетически омнипотентны. В связи с этим генетическая программа развития многоклеточного организма не является централизованной: в этом случае централизация собственной программы развития организма обусловлена уже не на генетическом уровне, а на уровне межклеточных взаимодействий, в результате которых осуществляется дифференцированная экспрессия генов отдельных клеток и тем самым достигается их неодинаковое развитие, необходимое для обеспечения функционально-структурной целостности организма как самостоятельной живой системы.
Все эти организмы — многоядерные одноклеточные, ценоцитные и многоклеточные — филогенетически возникли из монобионтов на основе экологически обусловленной прогрессирующей незавершенности процесса деления клеток, в результате чего после завершения репликации и сегрегации генетического материала деление клеток на том или ином этапе приостанавливалось.
В норме деление монобионтной клетки приводит к появлению двух дочерних клеток и в конечном итоге ведет к возникновению тех или иных ассоциаций монобионтов. Если дочерние клетки после их обособления расходятся, т. е. перестают контактировать, то в результате многократных делений возникает популяция пространственно разобщенных клеток. Если же дочерние клетки после их обособления сохраняют какие-либо тактильные контакты, т. е. окончательно не расходятся, возникает колония клеток, в которой все клетки структурно хотя и связаны, но физиологически вполне самостоятельны и в случае отделения от колонии, хотя бы под действием механических причин, продолжают нормально развиваться, образуя новые колонии.
Если процесс деления монобионтной клетки на той или иной стадии почему-либо приостанавливается, образуется агрегат, содержащий более чем один ядерный эквивалент, а в результате многократного повторения таких незавершенных делений возникает сложный многоядерный агрегат, структура которого может быть существенно различной в зависимости от того, на каком именно этапе прекращается процесс деления клеток. В этом смысле возможны три принципиально различных варианта.
Во-первых, если процесс деления ядерного эквивалента вообще не сопровождается делением клетки, формируются двухъядерные или многоядерные одноклеточные формы, такие, как рассмотренные выше Opalinina.
Во-вторых, если процесс клеточного деления приостанавливается после завершения формирования ядерно-цитоплазменных комплексов, еще не ограниченных обычными клеточными оболочками, возникают разного рода ценоцитные организмы — плазмодиальные, мицелиаль-ные или талломические. В этом смысле интересный пример представляют собою некоторые виды зигомицетов, относящиеся к группе муко-ровых: у них имеется не клеточный, т. е. ценоцитный, мицелий, в котором при старении образуются клеточные перегородки. Здесь мы видим, следовательно, как бы некоторую задержку в образовании клеточных перегородок, что явно указывает нам на тот путь, по которому шло развитие ценоцитности в филогенезе.
В-третьих, если клеточное деление завершается, но клетки не расходятся и сохраняют пространственный контакт, образуются многоклеточные агрегаты, которые могут быть интегрированы либо на уровне колонии, либо на уровне многоклеточной ор-ганизменной системы. В последнем случае агрегация монобионтов в структурно более сложную организменную систему закономерно приводит к их фукк зональной дифференциации, специализации и всесторонней коадап-тации в рамках этой новой системы и одновременно — к сужению их индивидуальной экологической валентности. В итоге всех этих изменений, на определенной стадии коадаптации монобионты полностью теряют свою организменную самостоятельность, т. е. исторически изначальная колония превращается в целостный, неделимый организм, находящийся на новом, более высоком уровне структурной агрегации. В этой новой — многоклеточной — организменной системе монобионты функционально низводятся до состояния внутренних структурных блоков.
Этот этап структурной агрегации обусловил, в частности, значительное усложнение информационной структуры организма благодаря 1) утрате им генетического моноцентризма и 2) вторичной централизации организменной информационной структуры на основе координирующих взаимодействий внутренних суборганизменных структурных блоков, в результате чего в этих блоках обеспечивалась дифференцированная трансляция генов соответствующих геномов.
В многоклеточном организме сложный комплекс межклеточных контактов представляет собою важнейший системообразующий фактор, определяющий целостность организменной системы и ее направленное развитие на тканевом, органном и организменном уровнях. Именно система межклеточных контактов придает совокупности контактирующих клеток новое качество — превращает их в тканевую систему и многоклеточный организм.
«Если при первичном контакте, клетки не отталкиваются, то наступает слипание за счет прочного соединения между ними, сопровождающееся образованием клеточных агрегатов. Это явление называется адгезией». Механизм действия межклеточных контактов сегодня еще далеко не вполне ясен, но имеющиеся данные уже позволяют «считать, что адгезионные свойства клеток зависят, прежде всего, от химического состава внешних примембранных слоев плазматических мембран... Теперь уже ни у кого нет сомнений в том, что именно плазматической мембране вместе с примембранными слоями и внутриклеточными мембранными образованиями принадлежит основная роль в обеспечении клеточных реакций и межклеточных взаимоотношений. Именно в ней заключена та специальная информация, которая обеспечивает узнавание, взаимодействие и реакцию на внешние воздействия».
Таким образом, информационная структура многоклеточных организмов, сравнительно с монобионтами, существенно усложняется за счет системы межклеточных контактов, которая, наряду с геномом, становится важнейшим координатором развития новой, многоклеточной организмен-ной системы.
Адаптивный смысл структурной агрегации монобионтов в более сложные организменные системы определяется многими факторами, из которых наиболее общее значение имеют три: 1) зависимость между интенсивностью метаболизма и размерами организма, 2) преимущества «блочной» конструкции тела и 3) взаимодействия организмов в системе хищник — жертва.
Отрицательная зависимость, связывающая относительную интенсивность метаболизма с размерами организма, показана для представителей самых различных систематических групп — от животных до микроорганизмов и высших растений, т. е. имеет общебиологический характер. Эта зависимость, имеющая крайне мало исключений, проявляется в том, что удельное потребление кислорода, т. е. его потребление в единицу времени в расчете на единицу массы тела, снижается с увеличением массы тела, причем этот процесс характеризуется определенным наклоном линий регрессии, который в среднем близок к 0,75.
До настоящего времени эта масштабная зависимость не получила исчерпывающего объяснения, однако можно предположить, что в основе ее лежит соотношение объемов и поверхностей тела, меняющееся с изменением его абсолютных размеров: чем крупнее организм и чем меньше, соответственно, его относительная поверхность, тем более облегчено, при прочих равных условиях, сохранение постоянства внутренней среды организма, поскольку ее принудительное, эктогенное изменение, бесспорно, пропорционально относительной величине поверхности организма, которая контактирует с агрессивной, т. е. дестабилизирующей, внешней средой. Таким образом, по представлениям автора, с увеличением размеров организма относительно облегчается гомеостаз, что и приводит к снижению относительной интенсивности метаболизма.
Происходящее с увеличением размеров, организма уменьшение относительной интенсивности метаболизма, означающее относительное биоэнергетическое «удешевление» жизненного процесса, бесспорно, служило мощным стимулом к увеличению размеров монобионтов. Однако увеличение размеров клетки экологически ограничено неизбежным уменьшением ее относительной поверхности и, следовательно, сокращением возг можностей адсотрофного питания, как и всех других жизненно важных процессов, связанных с адсорбцией или выделением тех или иных веществ через клеточную поверхность. Поэтому в процессе эволюционного увеличения размеров организма агрегация одноклеточных монобионтов явилась тем принципиально единственно возможным решающим шагом, который мог привести и действительно привел к существенному увеличению этих размеров и тем самым послужил одной из важнейших предпосылок к многоплановому разнообразию организмов и их широкому распространению в самых различных средах.
Таким образом, как способ разрешения противоречий между экологически целесообразным ростом размеров организма и уменьшением его относительной адсорбционной способности агрегация одноклеточных монобионтов в более сложные, многоблочные организменные системы была закономерным и неизбежным элементом процесса органической эволюции.
В индивидуальном развитии клетки те же противоречия ограничивают процесс ее роста и приводят к ее делению, в результате которого в дочерней системе оптимальное соотношение поверхности и объема восстанавливается.
Преимущества «блочной» конструкции многоклеточных организменных систем, открывающие путь к их неограниченной адаптивной модификации на очень простой и надежной структурно-информационной основе, несомненно, служили мощным стимулом к структурной агрегации монобионтов. Именно биологическая клетка как первичный суборганизменный конструкционный блок с автономной и централизованной информационной структурой явилась основным и универсальным фундаментом, на котором развился неисчерпаемый в своем разнообразии мир-многоклеточных живых существ. Сохранив сравнительно малые абсолютные размеры и, соответственно, большую относительную поверхность, клетка была оптимальным адсотрофным конструкционным блоком при формировании любых экологически обусловленных вариантов организменных систем. При любых функционально-структурных параметрах многоклеточной организменной макросистемы клетка сохраняла адсотрофность, оптимальную, как мы видели, для малых живых систем, и не препятствовала тем самым формированию сколь угодно крупных многоклеточных организменных макросистем, поскольку их интегральный обмен определялся жизнедеятельностью множества мелких и, вследствие этого, оптимально организованных адсотрофных подсистем.
Монобионтная структура тела оказалась, таким образом, лишь первым шагом живой природы, и шаг этот по необходимости был ограничен «минимальным» характером моноструктурной организменности. Монобионты дали бурную вспышку физиолого-биохимических вариаций и доказали свою эволюционную жизнеспособность на основе недостижимой для любых других организмов частоты смены поколений и обусловленного этой частотой высокого уровня изменчивости, чем обеспечили себе столь же высокий темп эволюции. Но при этом монобионты были консервативны в главном — они остались в рамках первичной моноструктурной организации, эволюционные возможности которой принципиально ограничены самой ее сутью: в любом случае эволюция замыкалась кругом относительно несущественных структурных изменений в рамках той же самой, элементарной, одноблочной конструкции со всеми ее лимитирующими особенностями, обусловленными соотношениями величин поверхностей и объемов неделимого тела.
Только агрегация монобионтов в более сложную, многоблочную организменную систему смогла разорвать этот замкнутый круг моноструктурности и привела к принципиальному расширению спектра адаптивной эволюции на основе многоклеточной конструкции тела.
Воздействие пресса хищников на живые объекты, находящиеся в составе агрегата, является статистически менее выраженным, чем на аналогичные объекты, пространственно обособленные и рассеянные в среде. Это способствовало как возникновению разного рода колоний прикрепленных организмов, так и структурной агрегации монобионтов в организменные системы более высокого структурного ранга.
Наряду с этим структурная агрегация монобионтов экологически стимулировалась и некоторыми другими аспектами воздействия хищников, и прежде всего тем, что по своим линейным размерам агрегаты монобионтов всегда превосходят тех отдельных монобионтов, которыми они образованы. Вследствие этого, по сравнению с отдельными монобионтами, их агрегаты и, в частности, образуемые ими организменные системы более высокого структурного ранга всегда находятся под воздействием более слабого пресса хищников, что, несомненно, также оказывает мощное стимулирующее воздействие на процесс структурной агрегации живых систем.
С увеличением абсолютных размеров организм как жертва становится доступным все более узкому кругу хищников. Этот факт уже давно известен экологам и, в частности, послужил основанием к тому, чтобы рассматривать рост рыб как адаптацию, направленную на ослабление пресса хищников. В этой связи совершенно ясно, что структурная агрегация как способ уменьшения пресса хищников экологически наиболее целесообразна для самых мелких организмов и, следовательно, в первую очередь именно для монобионтов как мелких и мельчайших блоков жизни, что и привело во многих случаях к агрегации их в более крупные организменные системы. К этому следует добавить, что абсолютные размеры хищника и жертвы определяют их отношения двояким путем:
1)линейные размеры жертвы определяют, насколько она доступна для хищника как объект определенного размерного класса. Этот аспект имеет существенное значение в любой системе хищник — жертва;
2)если и хищник, и жертва способны к активной локомоции, то в их отношениях появляется новый элемент — соревнование в скорости поступательного движения. В условиях пелагиали, т. е. в толще воды, где, по-видимому, происходил процесс агрегации каких-то протерозойских монобионтов в первые многоклеточные организменные системы, результат подобных соревнований целиком определяется законами гидродинамики.
Поскольку способность к активному поступательному движению свойственна не только одноклеточным животным, но и очень многим одноклеточным планктонным водорослям и бактериям, в планктонных системах хищник — жертва соревнование в скорости плавания представляет собою фактор, имеющий очень общее экологическое значение.
Закономерности поступательного движения в сплошных средах в связи с процессом экоморфогенеза детально рассматриваются в гл. 8; здесь же мы отметим, что при длине плавающих организмов менее 4,5 м доступные для них скорости плавания связаны с их линейными размерами прямой зависимостью. Это значит, что в диапазоне длин, характерных для монобионтов и их первичных многоклеточных агрегатов, доступные для организмов скорости плавания тем выше, чем больше их линейные размеры. Поэтому увеличение линейных размеров жертвы при прочих равных условиях означает и увеличение скорости ее плавания, а отсюда — и уменьшение ее доступности для хищника, что также, несомненно, явилось одной из экологических причин, стимулировавших процесс структурной агрегации монобионтов в более крупные организменные системы.
Поскольку все многоядерные одноклеточные, ценоцитные и многоклеточные организмы возникли из первичных организмов — монобионтов, мы называем их метабионтами, а их собственную программу развития — метабионтной. Использование основы цеха, в этих терминах имеет тот же смысл, что и в словах Meta-zoa и Metaphyta. К метабионтам относятся, следовательно, многоядерные одноклеточные, ценоцитные и многоклеточные организмы.
Таким образом, подобно тому как возникновение монобионтов было результатом структурной агрегации определенных доорганизменных объектов молекулярного уровня, так становление метабионтов представляет собою результат агрегации монобионтов.
Второй организменные уровень функциональной организации живой системы является, следовательно, результатом нового витка структурной агрегации, в ходе которого функционально-структурная и информационная организация живой системы принципиально усложнилась. При этом моноцентризм генетической информационной системы сохранился только на своем прежнем — монобионтной — уровне, т. е. на уровне клетки; централизация же собственной программы развития многоклеточного организма получила выражение на более высоком структурном уровне межклеточных взаимодействий.
Отсутствие централизованной в масштабах организма-метабионта генетической программы на первой стадии многоклеточности у таких организмов, как губки, приводит к большой автономии любых фрагментов тела и высокой регенеративной способности организма. По мере усложнения многоклеточной организменной системы программа онтогенеза все более централизуется на основе прогрессивного развития целостной системы морфопоэтических факторов, определяющей различную экспрессию генов в отдельных клетках В результате этого функциональная автономия отдельных фрагментов организма и его регенеративная способность уменьшаются, тогда как его функциональная компартментализация растет, т. е. повторяется цикл, уже пройденный в рамках одноклеточной структуры тела, филогенетически приведший к высококом-партментализированной конструкции тела, характерной для таких наиболее продвинутых групп одноклеточных, как жгутиконосцы или инфузории.
В эволюции метабионтов ясно выделяются два структурно различных пути: 1) адаптивное развитие в рамках метабионтной организменной структуры, что в громадном большинстве случаев оказалось экологически наиболее целесообразным и в некоторых систематических группах привело к появлению организмов наивысшей функционально-структурной сложности; 2) интегративное развитие колоний метабионтов, сопровождавшееся более или менее глубокой функционально-структурной специализацией и интеграцией образующих колонию организмов и прогрессирующей незавершенностью их вегетативного деления,
Обычное вегетативное размножение в конечном итоге завершается пространственным отделением дочернего организма от родительского, в результате чего в случае прикрепленных организмов возникает колония, т. е. некоторая ассоциация расположенных поблизости друг от друга коадаптированных структурно самостоятельных организмов, примерами чего могут служить хотя бы такие ассоциации, как клоны земляники или валлиснерии.
При незавершенном вегетативном делении зачаток дочернего организма сохраняет функционально-структурную связь с родительским организмом и в силу этого остается его частью. В этом случае при неоднократном вегетативном делении совокупность родительского и дочерних организмов превращается в «псевдоколонию» — сложную систему организменного уровня, в которой зачатки отдельных дочерних организмов могут иметь разную степень функционально-структурной самостоятельности, но всегда сохраняют ту или иную, онтогенетически исходную функционально-структурную общность на основе общесистемного обеспечения определенных жизненно важных функций.
Таким образом, структурная агрегация метабионтов имела в своей основе развитие процесса прогрессирующей незавершенности вегетативного размножения почкованием, в результате чего возникали узкоадаптированные гроздеподобные организменные системы, приспособленные преимущественно к прикрепленному, реже планктонному существованию. Функционально-структурная и информационная организация их оказалась агрегатно еще более сложной, чем в случае метабионтов. В частности, организменный уровень живой системы оказался еще более удаленным от молекулярного уровня материализации генетической программы развития системы, что, вероятно, явилось одним из факторов, обусловивших весьма ограниченное распространение организменной системы этого типа в органическом мире.
Как и в случае структурной агрегации монобионтов, агрегация мета-бионтов привела к их функциональной дифференциации, специализации и всесторонней коадаптации. В результате этой интегративной эволюции образующие колонию организмы-метабионты исторически утратили. свою организменную самостоятельность и их положение в колонии было низведено до уровня органов, а колония, напротив, приобрела черты функционально неделимой организменной системы, перейдя, таким образом, на новый, более высокий организменный уровень функциональной организации. На этом уровне метабионты стали суборганизменными структурными блоками, т.е. подсистемами нового организма, и, следовательно, заняли в новой организменной системе то самое положение, которое моно-бионты занимают в организменной структуре метабионтов. Поскольку все организмы этого нового, более высокого структурного уровня исторически возникли в результате интегративной эволюции тех или иных ассоциаций метабионтов, мы называем их ценометабионтами.
Если метабионтная конструкция организма оказалась в равной мере доступной грибам, растениям и животным, то ценометабионты представлены почти исключительно животными, из числа которых к ним относятся живые системы, традиционно называемые «колониями». Из вымерших форм таковы, в частности, некоторые кораллы и граптолиты, а в современной фауне — «колониальные» гидроидные полипы, сифонофоры, «колониальные» коралловые полипы, сгибающиеся, мшанки, некоторые «колониальные» крыло-жаберные, сложные «колониальные» асцидии, огнетелки и боченочники. Все эти ценометабионтные системы по своей функциональной организации находятся на организменном уровне, что, по мнению автора, однозначно определяется наличием у них общесистемной компартментали-зации организменного типа, т. е. наличием тех или иных структурно обособленных общесистемных органов. Более детально организменный характер всех перечисленных ценометабионтных систем обсуждается в п. 3.3; здесь мы лишь отметим, что интеграция функций в рамках общей организменной системы у них хотя и выражена в различной степени, но во всех случаях настолько глубока, что, бесспорно, доказывает их организменный характер.
У растений и грибов, при неограниченном росте и многократном ветвлении их талломов, побегово-корневых систем и мицелиев, отдельные фрагменты вегетативного тела, как правило, представляют собою обычные структурные блоки организма, не предназначенные для вегетативного размножения, в силу чего организменная целостность всей разветвленной системы не подлежит сомнению. И растениям, и грибам свойственны весьма разнообразные способы вегетативного размножения путем пространственного отделения от родительского организма тех или иных фрагментов, т. е. дочерних организмов. Не останавливаясь на описании этих способов, хорошо известных в ботанике и микологии, отметим лишь, что все они могут быть разделены на две группы.
Одна группа объединяет такие способы вегетативного размножения, которые основаны на простой фрагментации вегетативного тела, примером чего может служить фрагментирование мицелия грибов, состоящее в отделении от общей мицелиальной системы тех или иных неспециализированных фрагментов либо слабо специализированных фрагментов, каковы мелкие фрагменты мицелия — артроспоры.
Другая группа объединяет способы вегетативного размножения, основанные на отделении от родительского организма специализированных фрагментов. К их числу принадлежат, в частности, широко известные у растений выводковые почки, возникающие на поверхности растения и отпадающие на некотором этапе своего развития. Прорастание выводковых почек и развитие побегов дочерних растений начинается обычно еще до отпадения выводковой почки от родительского растения, как это происходит, например, у некоторых папоротников и толстянковых. В этом случае создается, следовательно, система, внешне аналогичная ценометабионтной конструкции, однако от перечисленных выше зоологических систем такого уровня она принципиально отличается своим временным характером. Родительское растение с находящимися на нем выводковыми почками по своему функциональному состоянию подобно почкующейся метабионтной гидре, а не ценометабионтному гидроидному полипу: как выводковые почки, так и почки на гидре в онтогенезе представляют собою лишь временные состояния, длительность существования которых в таком качестве обусловлена только ритмом вегетативного размножения. Постоянство же видимой картины папоротника Asplenium с прорастающими выводковыми почками на листьях или почкующейся гидры является результатом квазистационарности процесса, т.е. определяется постоянной сменой отпадающих и вновь возникающих почек.
Таким образом, в рассмотренных случаях ценометабионтная организ-менная система фактически не возникает: совокупность родительского растения с находящимися на нем выводковыми почками развивается в онтогенезе не по интегративному пути, а по пути прогрессирующего функционально-структурного разобщения родительского и дочерних организмов, в чем и состоит принципиальное отличие подобных макросистем от настоящих зоологических ценометабионтных систем организменного типа. Точно так же обстоит дело и в любых других случаях вегетативного размножения растений и грибов: все эти случаи по той же причине не приводят к образованию ценометабионтных организменных систем.
Весьма своеобразным вариантом гетерогенной ценометабионтной системы являются лишайники. При бесспорно симбиоти-ческой структуре их талломов и столь же очевидной генетической обособленности микобионта и фикобионта, они в то же время имеют ряд особенностей, характерных для организменных ценометабионтных систем.
В этом смысле совершенно исключительный интерес представляет, по мнению автора, полная аналогия некоторых эволюционно прогрессивных способов вегетативного размножения, специфических для лишайников и типично ценометабионтных пелагических гидроидных полипов — сифонофор. Тем и другим свойствен способ вегетативного размножения, состоящий в отделении фрагментов тела, содержащих минимальный комплект функционально необходимых структур, совокупность которых может дать начало новому организму. У лишайников такие фрагменты представлены изидиями и соредиями, т. е. маленькими клубочками гиф микобионта с находящимися между ними немногими клетками фикобионта, причем клубочки эти либо покрыты специальным коровым слоем, либо не имеют его. У сифонофор имеются аналогичные, по функциональному назначению, образования — кормидии, представляющие собою структурно обособленные группы важнейших органов. В состав кормидия входит обычно один кормящий полип — гастрозоид, один пальпон, выполняющий, как полагают, выделительные функции, и два гонофора, несущих половую функцию.
Функциональная аналогия соредии, изидий и кормидиев, несомненно, подтверждает закономерный характер возникновения подобного способа размножения у ценометабионтов и в этом смысле имеет совершенно исключительное значение для правильной интерпретации агрегатной структуры ценометабионтных организменных систем: названные фрагменты — соредии, изидии и кормидии — представляют собою элементарные единицы новой ценометабионтной организменной структуры. Именно поэтому в рассмотренных случаях вегетативного размножения лишайников и сифонофор происходит отделение уже не отдельных метабионтных структур, филогенетически соответствующих первичным особям изначальной колонии, а функционально целостного комплекса функционально различных коадаптированных метабионтов, выполняющих функции конкретных органов. Этот факт служит, по мнению автора, одним из веских доказательств организменного уровня ценометабионтной системы как в случае лишайников, так и в случае сифонофор.
Строго говоря, не все лишайники формально могут быть отнесены к ленометабионтам, поскольку фикобионт чаще всего представлен не мета-бионтом, а монобионтом, т. е. цианобактериями или одноклеточными водорослями, и, соответственно, слоевище лишайника чаще всего имеет моно-метабионтную основу, а не мета-мета бионтную, как у рассмотренных выше зоологических ценометабионтных систем.
Только в более редких случаях в талломе лишайника фикобионт, как и микобионт, является многоклеточным, что имеет место, когда фикобион-том служит, например, какая-то зеленая нитчатая водоросль, в частности Trentepolia; только в подобных случаях лишайник формально является ценометабионтом. Тем не менее общие черты ценометабионтной системы третьего организменного уровня свойственны лишайникам и при наличии монобионтного фикобионта, что подтверждается хотя бы наличием у них и в этом случае изидий и соредий. Эти и другие вопросы, связанные с определением уровня функциональной организации лишайников, подробнее обсуждаются в п. 3.3.
Таким образом, ценометабионты представлены только животными и лишайниками, причем из животных ценометабионтная конструкция тела оказалась доступной только для сравнительно примитивных групп — кишечнополостных, сгибающихся, мшанок, полухордовых и оболочников. По своему происхождению ценометабионтные животные и лишацники принципиально различны. Ценометабионтные животные исторически возникли в результате прогрессирующей незавершенности процесса вегетативного деления метабионтов, за счет чего на основе структурной агрегации организменной системы было достигнуто экологически целесообразное усложнение ее функциональной структуры. В данном случае, следовательно, онтогенез исторически изменялся таким образом, что в результате почкования метабионтного организма вместо колонии структурно обособленных метабионтов стал возникать единый сложный цено-метабионтный организм. Лишайники же сформировались в результате совершенно иного процесса — прогрессирующего симбиотического сближения изначально экологически независимых живых систем — гриба и фикобионта, функцию которого в разных случаях выполняли цианобак-терии или водоросли.
Большинство ценометабионтных животных относится к числу прикрепленных форм. Исключение составляют лишь очень немногие ползающие кораллы и ползающие мшанки, а также пелагические сифонофоры, пиросомы и бочоночники. Несомненно, что ограниченность круга це-нометабионтов примитивными, преимущественно прикрепленными животными непосредственно связана с происхождением ценометабионтной организменной структуры путем видоизменения процесса вегетативного размножения, которое, как и прикрепленный образ жизни, у более высокоорганизованных групп животных не встречается. Именно поэтому, надо полагать, такие группы животных, как членистоногие и позвоночные, не дали ценометабионтных вариантов организменной структуры.
Во всех случаях возникновение ценометабионтной конструкции тела было связано, как мы видели, с усложнением его агрегатной структуры, что повлекло за собою и усложнение его информационной структуры. На первых стадиях ценометабионтности, как и на ранних этапах метабионт-ности, централизация организменной системы была относительно слабой.
Эту относительно раннюю стадию становления ценометабионтной орга-низменности у животных мы можем видеть на примере ценометабионтных гидроидных полипов, у которых еще отсутствует единая нервная система, хотя уже имеется единая гастральная полость, т. е. трофическая и транспортная функции уже централизованы. Примерами наиболее высокоорганизованных ценометабионтов могут служить их подвижные формы, в частности пелагические планктонные сифонофоры и ползающие коралловые полипы — морские перья; у тех •и других имеется уже единая нервная система. Высокая степень централизации ценометабионтной организменной системы характерна для ползающих мшанок и планктонных бочоночников.
С усилением функционально-структурной интеграции ценометабионтной организменной системы ее собственная программа развития становится более централизованной на основе усложнения взаимодействий ее структурных блоков, т. е. первичных метабионтов, образующих ценометабионт-ную систему. Генетическая программа развития ценометабионтов, как и в случае метабионтов, централизована лишь на монобионтной, т. е. клеточном уровне.
Третий организменный уровень функциональной организации живой системы, представленный ценометабионтами, является, как видно из изложенного, результатом очередного — третьего в истории биологической эволюции — этапа структурной агрегации, в ходе которого произошло новое усложнение функционально-структурной и информационной организации живой системы. В этом процессе следует особо отметить развитие новой — ценометабионтной компартментализации тела, основанной на глубокой специализации исходных метабионтов и фактическом превращении их в органы, выполняющие конкретные жизненно важные функции в масштабах ценометабионтной организменной системы. При этом централизация собственной программы развития живой системы получила выражение уже на межметабионтном уровне, вследствие чего метабионтные программы исходных организмов-метабионтов сохранили лишь подчиненное значение и в информационной структуре третьего организменного уровня стали коадаптированными подпрограммами.
Поскольку ценометабионтная организменная система в своем возникновении ограничена наличием определенного способа вегетативного размножения метабионтов и поскольку она экологически связана с прикрепленными или медленно плавающими планктонными вариантами экоморф, она не получила столь широкого распространения в органическом мире, как монобионтная и особенно метабионтная системы, а напротив, как всякая узкоадаптированная система оказалась целесообразной только в сравнительно немногих случаях и была реализована только в весьма ограниченном числе вариантов.
Таким образом, существуют три уровня функциональной организации организменных живых систем, представленные, соответственно, моно-бионтами, метабионтами и ценометабионтами.
Надорганизменные живые системы
Поскольку все надорганизменные живые системы представляют •собою те или иные ассоциации организмов, они подразделяются на три группы, соответственно уровням функциональной организации тех организмов, которыми они образованы: 1) живые системы первого надорганизменного уровня, представленные ассоциациями монобионтов; 2) живые системы второго надорганизменного уровня, образованные ассоциациями метабионтов и 3) живые системы третьего надорганизменного уровня, образованные ассоциациями ценометабионтов. Наряду с этими группами существует адекватная им четвертая группа, представленная гетерогенными ассоциациями, состоящими одновременно из представителей первых трех групп.
Первый надорганизменвый уровень функциональной организации живой системы соответствует различным ассоциациям монобионтов — их популяциям, колониям, группам и т. п. Функционально-структурная-целостность всех этих ассоциаций обеспечивается на уровне межорганиз-менных взаимодействий, в ходе которых, на основе генетических программ конкретных организмов и при воздействии комплекса факторов окружающей среды, в каждый данный момент определяется стратегия развития надорганизменной системы как целого. Программа развития надорганиз-менной системы формируется, следовательно, как интегральный результат взаимодействия монобионтов друг с другом и с окружающей их средой,, хорошим примером чего могут служить хотя бы популяции одноклеточных жгутиковых водорослей типа Dunaliella или Chlamydomonas, которые при определенных условиях размножаются вегетативно, а при изменении этих условий начинают копулировать по типу гологамии.
Второй надорганизменный уровень функциональной организации живой системы представлен ассоциациями метабионтов — их популяциями, колониями, семьями и т. п. Функционально-структурная целостность всех этих систем, как и в надорганизменных системах монобионтов, обеспечена на уровне межорганизменных взаимодействий, однако эти взаимодействия по своему характеру более разнообразны и у высокоорганизованных Metazoa достигают исключительно высокой сложности. По своей функциональной и информационной структуре метабионтные надорганизменные системы в целом значительно сложнее монобионтных.
Третий надорганизменный уровень функциональной организации живой системы представлен ассоциациями ценометабионтов — их популяциями, колониями й т. д. Как уже было отмечено выше, ценометабионтные организмы в зоологии обычно именуются «колониями», что, по представлениям автора, явно противоречит фактической стороне дела и является отражением устаревших традиционных концепций, далеких от системного подхода. В действительности же, как следует из п. 2.2, колониями являются не сами организмы-ценометабионты, такие, как кораллы или мшанки, а образуемые ими надорганизменные ассоциации.
Примером простейшей ассоциации ценометабионтов может служить поселение мшанок, системная целостность которого обусловлена, в частности, генеративными связями отдельных гермафродйтных ценометабионтных особей, которые в процессе полового размножения обмениваются сперматозоидами. В ряде случаев внутрипопуляционная коадаптация ценометабионтов привела к возникновению более сложных систем надорга-низменного уровня. Из них наибольшей системной сложностью отличаются колонии ценометабионтных кораллов — коралловые рифы, закономерная структура и упорядоченность развития которых общеизвестны. Системный характер кораллового рифа обусловлен сложной совокупностью иерархически организованных подсистем, т. е. живых систем различного структурного уровня, собственные программы развития которых объединяются, образуя единую интегральную программу развития рифа как целостной системы.
Надорганизменные системы смешанного состава, образованные теми или иными совокупностями монобионтов, метабионтов и ценометабионтов, представляют собою наиболее сложные гетерогенные ассоциации — экосистемы различного ранга, стратегия развития которых обусловливается многими факторами. Информационная структура таких систем исключительно сложна; она определяется, по существу, на уровне биогеоценоза, рассмотрение чего уже выходило бы за рамки задачи настоящей книги.
По генезису образующих систему особей надорганизменные живые системы естественно подразделяются на гомогенные и гетерогенные, сложенные соответственно особями одного или разных видов. По степени функционально-структурной специализации отдельных особей и функциональной целостности можно различать системы гомоморфные, которые образованы морфологически близкими или идентичными особями, и гетероморфные, сложенные особями, существенно различающимися по своей морфологии. Как те, так и другие по степени структурной упорядоченности делятся на структурно неупорядоченные и структурно упорядоченные, которые характеризуются соответственно отсутствием или наличием какой-либо определенной закономерности в расположении особей относительно друг друга. По длительности своего существования надорганизменные живые системы могут быть разделены на временные и постоянные. Время существования первых значительно меньше п р юда существования данной экоморфы в онтогенезе, тогда как у вторых оно приближается к длительности этого периода или даже превосходит ее, в силу чего вся или почти вся жизнь особи проходит в условиях надорганизмен-ной системы и экологически нормальное существование особей вне этой системы в большинстве случаев уже невозможно.
Все это приводит к большому разнообразию надорганизменных живых систем: от пролетной стаи журавлей и простейшей гомогенной популяции однохромосомных вирусов до колонии мадрепоровых кораллов и биосферы все их параметры существенно изменяются. Из всего этого разнообразия в настоящей книге, в связи с проблемой экоморфы, целесообразно рассмотреть только постоянные структурно упорядоченные надорганизменные живые системы, поскольку только в этом случае может возникать реальный вопрос о границах организма и соотношениях экОморфы с надорганизменной системой, тогда как во всех остальных случаях и границы организма, и содержание системы адаптации экоморфы достаточно очевидны, т. е. особенности экоморф практически целиком определяются развитием адаптации организменного уровня.
Среди рассмотренных аспектов классификации надорганизменных живых систем наиболее существенным представляется деление их на гомогенные и гетерогенные, поскольку главным образом именно оно определяет как общие возможности интегративной эволюции недорганизменной системы, так и соответственно меру изменения, в ходе этой эволюции, экоморф организмов. Для гомогенных надорганизменных живых систем существенно также деление их на гомоморфные и гетероморфные, отражающее степень функциональной и структурной специализации особей в системе и, в то же время, меру функциональной пространственной дифференциации системы и ее функциональной интеграции как неделимого целого. Именно в. этом плане соотношения организма и постоянных структурно упорядоченных надорганизменных живых систем более детально рассматриваются в п. 3.3 в связи с анализом понятия организма.
Монобионтеые, метабионтные и ценометабионтные живые системы. Развитие живых систем как функция их структурной агрегации
Как было показано в п. 2.2.3, одна и та же функциональная организация живой системы — организменная или надорганизменная — может быть достигнута при различном уровне ее структурной агрегации — монобионтной, метабионтном или ценометабионтной. В соответствии с этим существуют, как мы видели, три организменных и три надорганизменных уровня функциональной организации живых систем, каждый из которых обусловлен определенным уровнем их структурной агрегации. В эволюционном аспекте каждый уровень структурной агрегации представляет собою результат определенного цикла развития, на протяжении которого происходит акт агрегации существующих предбиологических или биологических систем в системы более высокого структурного уровня.
Три цикла структурной агрегации прослеживаются в эволюционном развитии живых систем и, соответственно, выделяются три различных в этом отношении типа живых систем — монобионтный, метабионтный и ценометабионтный, каждый из которых отличается от предыдущего более высоким уровнем структурной агрегации системы. Таким образом, струк
турная агрегация живых систем явилась стержнем их закономерного циклического развития и послужила основой периодических явлений в их эволюционном функционально-структурном усложнении. В схематической форме периодический характер эволюционного развития живых систем показан в виде периодической таблицы, которая объединяет все существующие варианты живых систем.
Монобионтные живые системы
Первый цикл структурной агрегации живых систем — монобионтг ный — по своему содержанию состоял в агрегации предбиологических доорганизменных объектов — макромолекул и простых надмолекулярных агрегатов, совокупность которых в среде составляла, как иногда говорят, «первичный бульон», т. е. некоторое множество пространственно разобщенных и взвешенных в воде объектов, из которых на статистически реальной основе могли образоваться первичные организмы. Интегративная эволюция тех или иных ассоциаций этих объектов, т. е. макромолекул и их простых агрегатов, закономерно привела к их агрегации, в результате которой возникли первые живые организмы. Это был «минимальный» вариант живой системы, представленный организмами-монобионтами, информационная структура которых, как видно из изложенного, неделима благодаря их генетическому моноцентризму.
Таким образом, первая фаза монобионтного цикла явилась последним • этапом предбиологической эволюции и завершилась образованием первых живых систем — организмов-монобионтов. Тем не менее эта первая фаза монобионтного цикла по своему содержанию вполне аналогична начальной фазе любого последующего цикла структурной агрегации, поскольку она, как уже сказано, была представлена ассоциациями пространственно разобщенных интегративно эволюционирующих объектов, что характерно и для начальной фазы любого последующего цикла. Специфика монобионтного цикла состоит, следовательно, лишь в том, что на его протяжении произошло становление первых живых систем на основе структурной агрегации неживых химически активных объектов молекулярного уровня.
Монобионтная эра биологической эволюции продолжалась около 2 млрд. лет. Ее начало, совпадающее с появлением первых организмов, отделено от нашего времени почти 4 млрд. лет, а конец определяется временем возникновения первых многоклеточных, которые, судя по их разнообразию в верхнепротерозойской биоте и отдельным указаниям на значительно более ранние находки, появились в нижнем протерозое, т. е., видимо, около 2 млрд. лет назад. В течение громадного промежутка времени, который разделяет эти два события, живые существа на нашей планете были представлены только монобионтами, эволюция которых в этот период шла по пути адаптивного усложнения организменной системы в рамках одноклеточной конструкции тела.
Метабионтные живые системы
Второй цикл структурной агрегации живых систем — метабионт-ный — состоял в агрегации монобионтов на основе интегративной эволюции их надорганизменных ассоциаций. В ходе эволюции этих ассоциаций, наряду с усложнением обычных внутрипопуляционных межорганизменных отношений монобионтов, в условиях их колониального развития наметилась экологически обусловленная тенденция к их агрегации в структурно более сложные организменные системы метабионтного уровня. В результате этой агрегации возникли организмы-метабионты, которые выгодно отличались от монобионтов своими биоэнергетическими и конструкционными особенностями и своей способностью более успешно противостоять прессу хищников и на этой основе бурно эволюционировали. Именно эволюция метабионтов явилась источником разнообразия современной земной биоты и привела к появлению наиболее сложных организмов.
Моно-метабионтная эра биологической эволюции, в течение которой органический мир на нашей планете был представлен только монобион-тами и метабионтами, была, судя по всему, весьма непродолжительной. Если первые метабионты возникли, видимо, в нижнем протерозое, то уже в верхнем протерозое, на рубеже венда и рифея, присутствуют достаточно разнообразные ранние ценометабионты, в частности мягкие ценометабионтные кораллы, возникновение которых относится, следовательно, к более раннему периоду. Поэтому длительность моно-мета-бионтной эры измеряется, возможно, всего немногими сотнями миллионов лет.
Ценометабионтные живые системы
Третий цикл структурной агрегации живых систем — ценометаби-онтный — состоял в агрегации метабионтов. Интегративная эволюция колоний метабионтов под действием тех же экологических причин, что и в колониях монобионтов, в ряде случаев привела к агрегации организмов-метабионтов в более сложные организменные агрегаты, в результате чего в верхнем протерозое возникли организмы-ценометабионты.,
Моно-мета-ценометабионтная эра биологической эволюции, которая характеризуется присутствием в земной биоте одновременно всех трех структурно различных групп организмов — монобионтов, метабионтов и ценометабионтов, началась в верхнем протерозое и продолжается до настоящего времени.
Представляя собою конкретную адаптацию животных к прикрепленному образу жизни, ценометабионтная конструкция тела была в то же время лишена некоторых важных преимуществ метабионтной конструкции, и прежде всего свойственной метабионтам высокой функционально-структурной пластичности. Это и определило общее направление эволюции ценометабионтов, экологически ограничив его формированием прикрепленных и некоторых планктонных экоморф. И - если метабионты по своему качественному разнообразию заняли господствующее положение в земной биоте и их эволюция привела к появлению наиболее высокоорганизованных форм жизни, то ценометабионты остановились в своем развитии на весьма примитивном уровне общей организации.
Появление ценометабионтов сравнительно мало повлияло на общий ход биологической эволюции и не привело к каким-либо существенным переменам в биосфере. Если не считать развития колоний ценометабионтных рифообразующих кораллов, деятельность которых в Мировом океане, бесспорно, сделалась глобальным биогеохимическим фактором, то в остальном возникновение ценометабионтов, образно говоря, «осталось незамеченным»: лицо земной биоты и все важнейшие элементы ее интегрального жизненного баланса но-прежнему, как и в моно-метабионтную эру, всецело определяла и определяет совокупность монобионтов и метабионтов.
До настоящего времени в ряду циклического усложнения структурной агрегации живых систем третий — ценометабионтный — цикл является последним, в чем, видимо, находит отражение относительный характер адаптивной ценности агрегатного усложнения живых систем.
Адаптивная ценность процесса структурной агрегации живых систем
Адаптивная ценность процесса структурной агрегации живых систем достигает своего максимума при переходе от монобионтов к метабионтам, т. е. на метабионтном уровне, сочетающем высокую функционально-структурную пластичность организменной системы с относительной простотой и надежностью ее информационного обеспечения. В ходе дальнейшей агрегации происходят три процесса, ведущие к уменьшению ее адаптивной ценности.
Во-первых, резко увеличиваются линейные размеры конструкционных блоков* и вследствие этого столь же резко снижается их функционально-структурная пластичность, а следовательно, и их универсальность как конструкционного материала. Если клетка по своим функционально-структурным свойствам представляет собою действительно универсальный элементарный конструкционный блок и тем самым делает возможным по-истинне безграничное разнообразие многоклеточных организмов-метабион-тов, то строительный блок в виде недоразвитого многоклеточного мета-бионта со всеми содержащимися в нем тканями и зачатками органов, бесспорно, является значительно более громоздким, менее пластичным и узкоадаптированным, что с самого начала предопределяет его ограниченную пригодность как конструкционного материала. Понятно, что уже только, поэтому ценометабионты не смогли эволюционировать столь успешно, как метабионты.
Во-вторых, ухудшается информационная структура конструкционных блоков. Информационное обеспечение клетки, содержащей собственный геном, представляет собою оптимальную информационную структуру конструкционного блока, поскольку он изначально заключает в себе стандартную программу, которая может быть модифицирована, как мы видели, соответственно конкретной стратегии развития отдельных фрагментов метабионтного организма. Это обстоятельство создает клетке как конструкционному блоку второе важнейшее преимущество. Конструкционный блок в виде недоразвитого многоклеточного зачатка метабионта и в этом отношении, сравнительно с клеткой, бесспорно, проигрывает: его информационная структура оказывается несравненно менее гибкой, а главное — менее универсальной, чем в случае клетки. По своему макроструктурному уровню многоклеточный конструкционный блок значительно более, чем клетка, удален от молекулярного уровня материализации своей генетической программы развития, что, несомненно, осложняет информационное обеспечение такого многоклеточного блока и существенно ограничивает возможности адаптивной эволюции ценометабионтных организмов, по сравнению с метабионтными.
В-третьих, резко увеличиваются размеры организма, что при прочих равных условиях способствует, как мы видели, ослаблению пресса хищников и тем самым уменьшает стимул к дальнейшей структурной агрегации.
Ценометабионтные организменные системы как конструкционные блоки по тем же причинам пригодны еще менее, чем метабионтные, что и определило, судя по всему, отсутствие организмов еще более высокого структурного уровня, которые могли бы появиться в результате структурной агрегации ценометабионтов. Иначе говоря, те рассмотренные выше экологические стимулы, которые способствовали агрегации монобионтов, на уровне метабионтов значительно уменьшились, а на уровне ценометабионтов исчезли вовсе, в связи с чем прекратился и дальнейший процесс структурной агрегации живого, завершившийся появлением ценометабионтных систем.
При этом продолжается, однако, ничем не ограниченный во времени, грандиозный в своей многоплановости внутрициклический процесс адаптивной эволюции живых систем. В современных земных условиях процесс этот ограничен лишь тремя конкретными уровнями структурной агрегации организмов — монобионтным, метабионтным и ценометабионтным.
Автобионтные и анавтобионтные живые системы
Деление живых систем на автобионтные и анавтобионтные отражает два принципиально различных способа организации процессов метаболизма. Как процесс обмена веществом и энергией между организмом и окружающей его средой метаболизм может быть организован двумя существенно различными способами, в основе которых лежат принципиально различные экоморфологические предпосылки.
Первый способ — автобионтный — заключается в осуществлении всего комплекса обменных процессов собственными силами организма. Центральным, важнейшим видоспецифическим звеном этого метаболического комплекса являются репликация генома и синтез белка. Некоторые другие, функционально не менее важные, но менее видоспецифические процессы, прежде всего связанные с накоплением энергии в форме адено-зинтрифосфата, в отдельных случаях могут осуществляться на основе использования необходимой материальной базы другого организма. Это, в частности, можно видеть на примере бактерий из группы хламидий, которые, будучи облигатными внутриклеточными паразитами, приспособились к существованию в среде, богатой АТФ, и поэтому практически утратили способность к самостоятельному синтезу АТФ, за что их называют, «энергетическими паразитами». Тем не менее даже и в этом крайнем случае — при использовании экзогенных источников АТФ — репликация генетического материала и синтез белка осуществляются собственными силами организма, в чем и состоит характернейшая черта рассматриваемого способа организации метаболических процессов. Ясно, что для этого организм должен располагать, соответственно, собственными ферментными и белоксинте-зирующими аппаратами. Из этого, в свою очередь, вытекают два следствия:
1)наличие указанных ферментных и синтезирующих аппаратов возможно только на основе определенного комплекса морфологических адаптации на уровне макромолекул и надмолекулярных агрегатов различной сложности, развитие которых делает организм в структурном отношении достаточно сложным;
2)при таком способе организации обменных процессов организм неизбежно должен иметь многоплановые прямые связи с внешней средой, в том числе трофические, обеспечивая как поступление вещества и энергии извне, так и удаление возникающих продуктов обмена в окружающее пространство.
Этот способ организации процессов метаболизма свойствен всем клеточным организмам, т. е. все клеточные организмы — а в т о б и о н т ы. Кроме того, автобионтами были, несомненно, и все первичные доклеточные организмы, поскольку они могли возникнуть не иначе как живые системы, самостоятельные в трофоэнерге-тическом отношении. В этой связи важно еще раз подчеркнуть, что понятие автобионтности связано не с клеточной структурой организма как таковой, а с определенным способом организации метаболических процессов, хотя в современной земной биоте автобионты представлены только клеточными организмами.
Второй способ — анавтобионтный,— в отличие от первого, состоит в осуществлении всех обменных процессов на основе обязательного использования энергозапасающих, ферментных и синтезирующих аппаратов другого организма, что в принципе возможно только для облигатных внутриклеточных паразитов, поскольку все эти аппараты представляют собою структуры субклеточного уровня. Из этого вытекают два следствия:
1)такой способ организации метаболических процессов открывает широкие возможности к морфологическому упрощению организма, так как последний лишен необходимости иметь собственные энергозапасающие, ферментные и синтезирующие аппараты;
2)в этом случае организм не имеет прямых трофических и энергетических связей с окружающей его средой, поскольку не только поступление вещества и энергии извне, но и все синтезы обеспечиваются организмом хозяина.
Такой способ организации процессов метаболизма свойствен всем вирусам, т. е. все вирусы — анавтобионты. Суть паразитирования вирусов состоит в том, что проникающая в клетку вирусная нуклеиновая кислота навязывает клетке свою программу синтеза, в результате чего синтезирующий аппарат клетки вместо специфических для клетки синтезов начинает осуществлять синтезы вирусспецифические. На внутриклеточной стадии своего развития вирус представляет собою лишь подсистему в целостной живой системе вирус — клетка, выполняя в этой системе функции информационного центра, тогда как метаболические функции в масштабах системы целиком лежат на клетке. В соответствии с этим вирус не питается в обычном понимании этого термина и не растет: в инфицированной клетке под действием вирусной нуклеиновой кислоты происходит лишь синтез нового поколения вирионов, обеспечиваемый материальными, энергетическими и синтезирующими средствами клетки.
Таким образом, рассмотренные способы организации процессов метаболизма имеют прямо противоположные экоморфологические следствия, которые создают самые фундаментальные различия между организмами. Эти различия касаются самой основы важнейших жизненных процессов и по своей глубине и функциональной многозначности, бесспорно, превосходят все другие различия между формами живой природы.
Автобионтные живые системы
К числу автобионтных живых систем принадлежат, следовательно, все древнейшие доклеточные и все клеточные организмы, а также все образуемые ими надорганизменные ассоциации любого ранга.
По качественному функционально-структурному разнообразию автобионтные системы в земной биоте составляют и, несомненно, всегда составляли громадное большинство живых систем, что определяется их трофоэнергетической самостоятельностью и связанной с нею широчайшей адаптивной вариабельностью, которая создает основу для их многоплановых эволюционных изменений. Все многообразие земной жизни, как существующей сегодня, так и существовавшей в прежние геологические эпохи, практически почти целиком обусловлено именно адаптиввой эволюцией автобионтов. Аспекты разнообразия автобионтов детально рассматриваются в гл. 7. Здесь мы лишь отметим, что это разнообразие в своих наиболее крупных элементах определяется трофическими факторами.
Совокупность организмов-автобионтов в единой экоморфологической системе организмов. образует один из двух таксонов самого высокого ранга — царство автобион.
Анавтобионтные живые системы
Анавтобионтные живые; системы представлены только вирусами и их ассоциациями. Они представляют собою пример крайней минимизации живой системы, что определяется их трофоэнергетической несамостоятельностью, т. е. полной метаболической зависимостью от организма-хозяина. Эти свойства анавтобионтных живых систем приводят к их сравнительно узкой адаптированности и функционально-структурному однообразию: будучи неподражаемо разнообразными в малом и образуя множество мелких физиолого-биохимических вариаций различного плана, они, в то же время, удивительно однотипны в части своих основных функциональных и структурных особенностей. Поэтому если автобион составляет основу качественного разнообразия земной биоты, то вклад анавтобиона в это разнообразие сравнительно невелик.
Совокупность организмов-анавтобионтов в единой экоморфологиче-ской системе организмов образует самостоятельный таксон высшего ранга — царство анавтобион.
Анавтобион принципиально отличается от автобиона по своему генезису. Если автобион, т.е. совокупность клеточных организмов, представляет собою бесспорно монофилетическое царство, то анавтобион столь же бесспорно полифилетичен: по общему убеждению, разные группы вирусов происходят от организмов из самых различных систематических групп, представляя, видимо, «побочный продукт» их филогенеза.
Принадлежность вирусов к числу организмов и даже к числу живых систем в настоящее время не является общепризнанной, что вынуждает нас остановиться на рассмотрении этих вопросов в пп. 2.2.5.4 и 3.2.5.
Вирус как живая система
Глубокие различия между клеточными организмами и вирусами привели к разным толкованиям природы вирусов. Если живая природа любых клеточных организмов ни у кого не вызывает сомнения, то на вопрос о том, являются ли живыми вирусы, различные исследователи до настоящего времени отвечают по-разному. После того как Ф. Боуден и Н. Пири получили истинные трехмерные кристаллы вирионов вируса кустистой карликовости томатов, этот вопрос сделался предметом оживленной дискуссии, не давшей, однако, однозначных результатов, поскольку, как правильно заметили А. Гиббс и Б. Харрисон, «ответ зависит от того, какой критерий положить в основу определения жизни». В экоморфологии эта проблема приобретает особое значение в связи с тем, что соотношения клеточных и неклеточных форм жизни отражают многие принципиально важные аспекты развития экоморф, благодаря чему вирусологический материал составляет неотъемлемый элемент общей картины экоморфогенеза.
В основе дискуссий о том, являются ли вирусы живыми, лежат два момента: 1) биологическая «несамостоятельность» вирусов, их неспособность размножаться без использования энергозапасающих, ферментных и синтезирующих аппаратов живой клетки и 2) их структурная простота, подчас, казалось бы, несопоставимая с теми привычными для нас канонами, которые были восприняты нами при изучении клеточных организмов. Эти обстоятельства в основном и служат причиной того, что некоторые вирусологи до настоящего времени отказывают вирусам в праве называться живыми.
Характерным примером подобной концепции может служить точка зрения П. Зенгбуша: «Являются ли вирусы живыми? На этот вопрос можно с уверенностью ответить отрицательно... вирусы нельзя считать живыми существами. Они не могут самостоятельно размножаться, но
Рис. 2
Кристаллы, образованные агрегатами вирионов вируса кустистой карликовости томатов. X 250. По Bawden, Pirie.
они заставляют размножать себя и, таким образом, являются паразитами на генетическом уровне». Как видим, это высказывание противоречиво: с одной стороны, оно отрицает принадлежность вирусов к живым существам, с другой — признает паразитизм вирусов, т. е. наделяет их свойством, присущим исключительно организмам, поскольку паразитизмом принято называть определенного рода отношения между организмами.
Суть дела, однако, даже не в формальном несоответствии этих двух утверждений в цитированном тексте, а в том, что вся совокупность известных к настоящему времени фактических сведений о вирусах уже не позволяет сомневаться в их живой природе. Именно поэтому, пытаясь избежать признания живой природы вирусов, Зенгбуш снова вынужден занимать противоречивую позицию, когда определяет свойства живых систем и свойства вирусов. «Каждая живая система, существующая сегодня, содержит белки, обладающие свойствами катализаторов, и нуклеиновые кислоты, играющие роль носителей информации... Ни белки, ни нуклеиновые кислоты по отдельности не могут развиться в живые системы. Лишь взаимодействие между этими двумя классами молекул приводит к возникновению свойств, характерных для живой системы». В то же время, Зенгбуш определяет вирусы как «инфекционные единицы, состоящие из нуклеиновой кислоты и белка», т. е. указывает именно те их свойства, которые, по его мнению, характерны для живых систем.
Утверждение Зенгбуша, «что вирусы есть не что иное, как определенные формы состояния генетической информации... которые при случае могут обратимо перейти в другое состояние», также не может служить дополнительным обоснованием его тезиса, что «вирусы нельзя считать живыми существами». Достаточно напомнить, что любой организм характеризуется «определенным состоянием генетической информации, которое при случае обратимо переходит в другое состояние», примером чего могут служить хотя бы диплоидные и гаплоидные фазы в ядерном цикле эукариот.
Аналогичная точка зрения, отрицающая живую природу вирусов, принадлежит Е. Гюнтер с соавторами, которые определяют вирусы как объекты, «состоящие из нуклеиновых кислот и белков, но не являющиеся живыми», что несколько подробнее рассматривается в п. 2.4 в связи с определением живой системы, где отмечается непоследовательность подобной позиции.
Таким образом, отрицание живой природы вирусов закономерно входит в противоречие с общеизвестными фактами их функционально-структурной организации, которые свидетельствуют о наличии важнейших фундаментальных аналогий между вирусами и клеточными организмами.
Большинство современных исследователей признает живую природу вирусов на том основании, что они «в большей или меньшей степени обладают или, вернее, сохраняют основные атрибуты жизни, включая такой кардинальный атрибут, как способность к эволюции».
Вирусам, даже самым простым,-таким, как сателлиты, в полной мере свойственны основные свойства живого: 1) они имеют собственную программу развития в виде генома, принципиально аналогичного геномам клеточных живых систем; 2) они характеризуются определенной иерархической функционально-структурной организацией; 3) даже наиболее простые из них достаточно сложны, хотя бы уже потому, что состоят из тех же самых высо-косложных биополимеров — нуклеиновых кислот и белков, что и все другие известные нам организмы. Таким образом, сомневаться в живой природе вирусов в настоящее время уже нет оснований. «Вирусы — это живые существа, организация которых еще менее сложна, чем у бактерий... Вирусы относятся к облигатным внутриклеточным паразитам; однако характерной особенностью вирусов, отличающей их от других внутриклеточных паразитов, является то, что они паразитируют на генетическом уровне».
Кроме информации конкретно биологического характера, касающейся существования неклеточных живых систем, и помимо разработки очень важных в практическом отношении медицинских аспектов вирусологии, открытие и исследование вирусов принесло биологии фундаментальные результаты общенаучного, эвристического характера, открывающие возможность новой оценки многих известных фактов и существенно корректирующие наши представления о формах и возможностях развития живых систем, о границах живого и неживого, об организме, о соотношении организма и популяции и многом другом. Многие процессы, хорошо изученные для клеточных организмов, на примере вирусов предстают в новом свете, и эта «новая модель» жизни принципиально углубляет наши познания о сущности живого, в том числе и в вопросе о свойствах живых систем.