Матричний синтез білка
СОДЕРЖАНИЕ: Біосинтез білка. Будова рибосом прокаріотів та еукаріотів. Роль мембран у формуванні клітинних компартментнів. Ароморфози як біологічний процес. Асиметричність плазматичної і внутрішніх мембран клітини. Транспортування речовин через мембрани.Міністерство освіти та науки України
Житомирський державний технологічний університет
Кафедра природничих наук
Контрольна робота
з предмету:
«Біологія»
Виконав:
Перевірив:
Житомир
2010
Чому синтез білка називають матричним
Біосинтез білка – складний процес, який регулюється великою кількістю ферментів. Він потребує енергії та участі іонів-активаторів. Ми розглянемо лише деякі ключові моменти. Процес біосинтезу білка умовно можна поділити на такі етапи:
· Вихід зрілої мРНК з ядра у цитоплазму.
· Активація амінокислот.
· Утворення ініціюючого комплексу за участю мРНК і рибосоми та ініціація поліпептидного ланцюга.
· Елонгація – побудова поліпептидної послідовності.
· Термінація – закінчення синтезу поліпептидного ланцюга у вигляді первинної структури.
· Процесинг – утворення вторинної і третинної структур білка.
- Активація амінокислот – це приєднання карбоксильної групи амінокислоти до 3-кінця відповідної тРНК.
Амінокислота приєднується до такої тРНК, антикодон якої комплементарний генетичному коду. Цей процес проходить з використанням енергії. Сумарна реакція активації:
Амінокислота + АТФ + тРНК + Н2 О
Аміноацил-тРНК + АМФ + 2Фн
Цю реакцію каталізує група ферментів – аміноацилсинтетази. Для кожної амінокислоти є свій фермент. Сполуку, яка утворюється, називають за назвою відповідної амінокислоти з закінченням –ил (-іл). Наприклад, комплекс між амінокислотою метіоніном і метіоніновою тРНК називають метіоніл-тРНК, комплекс між лізином і лізиновою тРНК – лізил-тРНК тощо.
Ініціюючий комплекс – це система, яка забезпечує початок синтезу білка. У еукаріотів він утворюється у цитоплазмі або на поверхні шорсткого ендоплазматичного ретикулуму, шляхом з’єднання у єдину систему мРНК, рибосоми і аміноацил-тРНК. У прокаріотів утворення ініціюючого комплексу проходить лише у цитоплазмі.
- Ініціюючий комплекс складається зі стартової аміноацил-тРНК, рибосоми і зрілої мРНК.
Перша (стартова) аміноацил-тРНК, з якої починається утворення пептидного ланцюга, приєднується до стартового кодону мРНК. Як у прокаріотів, так і у еукаріотів стартовий кодон однаковий – AUG. Таблиця генетичного коду (табл. 1.24) показує, що такий кодон відповідає амінокислоті метіоніну. Але метіоніл-тРНК стартова аміноацил-тРНК лише у еукаріотів. У прокаріотів стартовою аміноацил-тРНК є особлива формілметіоніл-тРНК. Вона утворена нестандартною амінокислотою – формілметіоніном (рис. 1.41).
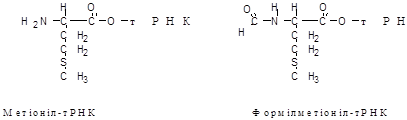
Рис. 1. Стартові аміноацил-тРНК
Рибосоми – це клітинні структури, які утворені з двох субодиниць (великої та малої). Вони не мають оболонок і складаються з рРНК і білка. Рибосоми прокаріотів і еукаріотів близькі за будовою (рис. 1.42). Кожна з них має дві спеціальні ділянки, які називають Р-ділянка (пептидильна) і А-ділянка (аміноацильна).
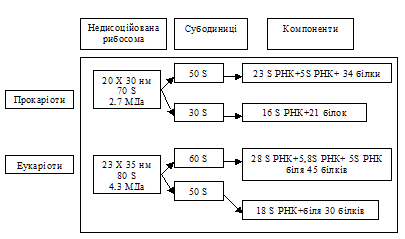
Рис. 2. Будова рибосом прокаріотів та еукаріотів
Розглянемо формування ініціюючого комплексу на прикладі прокаріотичної клітини. Воно поділяється на ряд послідовних дій:
· Спочатку формілМет-тРНК до Р-ділянки малої субодиниці рибосоми.
· Далі до Р-ділянки малої субодиниці рибосоми приєднується ініціююча частина мРНК, яка розташована на 5-кінці, на відстані не менше 25 нуклеотидів від початку молекули.
· Наступний заключний етап – приєднання великої субодиниці.
- Після утворення ініціюючого комплексу розпочинається синтез поліпептидного ланцюга, який має назву елонгації.
Наступна аміноацил-тРНК визначається за принципом комплементарності між кодоном і антикодоном. Вона приєднується до А-ділянки рибосоми.
- Під дією ферменту пептидилтрансферази між аміногрупою першої амінокислоти та карбоксильною групою другої амінокислоти утворюється пептидний зв’язок.
Важливою особливістю пептидилтрансферази є те, що цей фермент фіксований на рибосомі. Він постійно “прикріплений до місця роботи“. Після правильного приєднання аміноацил-тРНК і утворення пептидного зв’язку між амінокислотами відбувається транслокація.
- Транслокація – це зсув ініціюючого комплексу на три нуклеотиди вздовж молекули мРНК
Відомо, що різні білки починаються з різних амінокислот. Це вступає у протиріччя з тим фактом, що початковою аміноацил-тРНК завжди є формілметіонінова або метіонінова. Розв’язується ця проблема досить просто – ініціююча аміноацил-тРНК (формілметіонінова) не утворює пептидного зв’язку з наступною амінокислотою. Тобто, перша транслокація рибосоми відбувається не за правилами. Це, начебто, “холостий хід”.
- Інформація з мРНК зчитується у напрямку 5 3, а поліпептидний ланцюг росте у напрямку N C.
Закінчення процесу біосинтезу називають термінацією. Воно відбувається тоді, коли на мРНК зустрічається один з трьох стоп-кодонів: UАА, UАG, UGА.
Дозрівання поліпептидного ланцюга називається процесингом. Він полягає в утворенні третинної конформації молекули. Під час процесингу можливі вилучення деяких амінокислотних послідовностей. У складних білках процесинг включає приєднання небілкових груп тощо.
Процес біосинтезу білка – один з найскладніших у клітині. Не всі його нюанси на сьогодні відомі. Найбільш детально досліджений біосинтез білка прокаріотичного організму E. coli. Але навіть тут є ще деякі нез’ясовані моменти. Ми розглянули біосинтез білка лише схематично.
Роль мембран у формуванні клітинних компартментнів
Плазматичні мембрани і мембранний транспорт
Однією з найважливіших характеристик клітин є характеристика її оболонок або мембран. Саме завдяки цим структурам живі істоти відділяються від навколишнього середовища, створюючи якісно нову форму існування хімічних елементів у природі – живу речовину. Але еволюція еукаріотичних клітин не зупинилася лише на відділенні організму від середовища, а пішла далі. Еукаріоти мають не тільки зовнішню, але і внутрішні мембранні структури.
- Виникнення спеціальних оболонок всередині клітини було одним з вирішальних етапів у розвитку життя взагалі.
У еволюційній теорії такі зміни мають спеціальну назву – ароморфози.
- Ароморфоз – це такі еволюційні зміни, які ведуть до загального підвищення організації, збільшують інтенсивність життєдіяльності, але не є вузькими пристосуваннями до різко обмежених умов існування.
- Ідіоадаптація – це дрібніші еволюційні зміни організмів, які сприяють пристосуванню до певних умов життєвого середовища.
Ароморфоз завжди приводить до біологічного прогресу. Ці зміни не мають прямого пристосувального значення. Але ароморфози, підвищуючи інтенсивність життєдіяльності організмів, забезпечують відносну незалежність живих систем від довкілля.
Вивчення особливостей будови і функцій клітини закономірно розпочати з особливостей будови зовнішньої плазматичної мембрани.
- Плазматичною (або цитоплазматичною) називають мембрану, яка відділяє клітину від зовнішнього середовища.
Іноді зовнішню мембрану, на відміну від внутрішньої, називають плазмолемою (грец. рlasma – форма, lemma – оболонка). Плазматична мембрана характерна як для еукаріотів, так і для прокаріотів. У сучасних живих організмів вона виконує такі функції:
· забезпечує збереження відмінностей між клітинним вмістом і навколишнім середовищем. Це своєрідний бар’єр, який уможливив раннім формам життя відмежуватися від навколишнього середовища;
· визначає величину і форму клітин;
· це місце протікання енергетичних та синтетичних процесів;
· активний фільтр, який регулює обмін речовин між клітиною та довкіллям.
Як показали спеціальні дослідження, принципи побудови зовнішньої і внутрішніх мембран практично однакові. Всі наявні відмінності пов’язані виключно з функціональними особливостями конкретної мембранної структури.
Мембрани, які знаходяться всередині клітини і обмежують її компартменти, називають внутрішніми мембранами. Внутрішні мембрани за своєю будовою принципово подібні до плазмолеми. Їм властива здатність до самозбирання і самопідтримування цілісності, латеральна дифузія і асиметричність.
- Асиметричність плазматичної і внутрішніх мембран клітини приводить до того, що внутрішні порожнини ендоплазматичного ретикулуму, апарату Гольджі, порожнини мікротілець і лізосом топографічно відповідають зовнішній поверхні плазматичної мембрани клітини.
Внутрішні мембрани властиві лише еукаріотичним клітинам. Клітини прокаріотів мають лише одну плазматичну мембрану. Виникнення внутрішніх мембран – це один з важливих ароморфозів. Виникнення внутрішніх мембран пов’язане з розв’язанням проблеми співвідношення між об’ємом і площею поверхні у клітині, що важливо. Зростання площі поверхні значно відстає від зростання об’єму. Площа розраховується у квадратних (м2 ), а об’єм – у кубічних одиницях (м3 ). Площа поверхні клітини забезпечує обмін клітини з навколишнім середовищем, а об’єм клітини відповідає внутрішньому середовищу. Е. coli у 10 разів менша за найменшу еукаріотичну клітину. В середньому еукаріотичні клітини у 1000 разів більші за прокаріотичні. Це дає можливість прокаріотам обходитися однією плазматичною мембраною, яка іноді утворює випинання (мезосоми). Еукаріотам необхідні внутрішні мембрани. У еукаріотів площа внутрішніх мембран перевищує розміри плазмолеми. Наприклад, поверхня ендоплазматичного ретикулуму у гепатоцитах перевищує площу плазмолеми у 25 разів, а в ендокринних клітинах підшлункової залози ця різниця становить 12 разів.
Функції внутрішніх мембран подібні до функцій плазмолеми, але є і певні особливості. Внутрішня мембрана:
· це місце протікання і компонент метаболічних реакцій;
· вона розділяє різні, несумісні хімічні реакції;
· вона утворює місткості для зберігання метаболічних продуктів.
Мембрани – це не лише бар’єри між клітинами, але і місце протікання багатьох реакцій. Наприклад, перетворення енергії неможливе без участі внутрішніх мембран мітохондрій. Багато ферментів є мембранними білками, які без наявності мембрани втрачають свою активність або її частину. Наприклад, вже згадувана (Na+ -K+ )-АТФ-аза.
Компартменталізація уможливлює клітині просторове суміщення протилежних хімічних реакції. Наприклад, синтез жирних кислот проходить у цитозолі, а їх розщеплення – у мітохондріях.
Внутрішні мембрани служать для зберігання різних, іноді агресивних, речовин. Наприклад, лізосоми містять ферменти, здатні розщепити клітину. Саме мембрани захищають живу клітину від автолізису (самопошкодження).
Внутрішні частини еукаріотичних клітин (компартменти) умовно можна поділити на дві групи:
· компартменти, обмежені однією мембраною;
· компартменти, обмежені двома мембранами.
До першої групи відносять ендоплазматичний ретикулум, апарат Гольджі, лізосоми, пероксисоми (мікротільця), вакуолі, піноцитозні пухирці та інші.
Другу групу утворюють органели з подвійними мембранами – ядро, мітохондрії і хлоропласти. Ці компартменти мають не лише подвійну мембрану, але і містять у собі спадкову інформацію у формі ДНК. Виділення ядра як компартменту, оточеного подвійною мембраною, скоріше традиційний підхід, ніж реальна особливість будови. Як буде детально розглянуто у подальшому, ядерні мембрани морфологічно пов’язані з ендоплазматичним ретикулум. Подвійні мембрани мітохондрій та хлоропластів морфологічно не пов’язані з іншими мембранними структурами еукаріотичних клітин. Відносно походження цих компартментів зараз поширеною є теорія про клітинний симбіоз. Вважається, що мітохондрії і хлоропласти – це примітивні прокаріотичні організми, які об’єдналися з древніми еукаріотами для спільного взаємовигідного існування.
хімічний склад мембран
Відповідно до загальноприйнятої рідиннокристалічної моделі, за хімічним складом всі мембрани містять три класи органічних сполук:
· мембранні ліпіди;
· мембранні білки;
· мембранні вуглеводи.
Клітинні мембрани складаються з амфіпатичних ліпідів. У складі однієї молекули таких ліпідів одна або дві жирні кислоти етерифікуються гліцеролом або сфінгозином. У першому випадку утворюються гліцероліпіди, у другому – сфінголіпіди. До третього атома Карбону приєднується фосфат (фосфоліпіди) або цукор (гліколіпіди). Фосфат, у свою чергу, може зв’язуватися з іншою полярною групою – холіном. Крім ліпідів, які містять жирні кислоти, до складу мембран може входити також простий ліпід – холістерол, який має складну гетероциклічну “голівку”, яка може нести заряд, і вуглеводневий “хвіст”.
- Всі клітинні мембрани мають принципово однакову основу, а саме мембрани – це ліпідні подвійні шари, або бішари.
Утворення бішарів – це природна властивість фосфоліпідів та гліколіпідів у водному середовищі. Це найвигідніший, з енергетичного погляду, стан амфіпатичних молекул у даному середовищі.
- Утворення ліпідної основи біомембран – це процес, який проходить шляхом самозбирання.
Ліпідні бішари замикаються самі на себе, утворюючи закриті відсіки (компартменти). Внаслідок такої самозамкненості закриваються вільні краї, на яких гідрофобні “хвости” могли б стикатися з водою. З тієї ж причини компартменти, побудовані з ліпідних бішарів, здатні самі лікувати свої пошкодження. Попробуйте розділити на дві половини мильну бульбашку – ви отримаєте дві нові бульбашки, а не дві половинки однієї бульбашки. З цього випливає одна з найважливіших властивостей біомембран:
- Мембрани здатні не лише до самозбирання, але і до самозбереження біліпідного шару.
Дослідження, проведені на штучних та природних біомембранах, показали, що ліпіди у межах одного моношару постійно рухаються в одній площині з великою швидкістю – течуть. Така динамічність має назву латеральної дифузії. Таким чином, ще однією важливою рисою біомембран є те, що:
- Ліпідний бішар – це двомірна рідина, здатна до латеральної дифузії.
Рухливість ліпідів у бішарі має велике, багато у чому ще не зовсім з’ясоване, біологічне значення. Вона значною мірою зв’язана з мембранним ліпідом холістеролом. Швидкість латеральної дифузії залежить від температури. При певній температурі латеральна дифузія припиняється. Мембрана переходить з рідкого у кристалічний стан.
Ліпідний склад шару, який стикується з цитоплазмою клітини, принципово відрізняється від складу зовнішнього моношару, в якому знаходиться основна частина гліколіпідів. Для зовнішнього шару деяких клітин характерна також наявність холіну. Фосфоліпіди в основному характерні для внутрішніх шарів.
- Важлива риса ліпідного бішару плазматичної мембрани полягає у тому, що він асиметричний, тобто зовнішній та внутрішній моношари різні за хімічним складом.
Така асиметрія може мати велике значення в орієнтації мембран. Вона закладається в момент синтезу відповідних ліпідів. Різний ліпідний склад характерний не тільки для внутрішнього і зовнішнього моношарів ліпідних бішарів, але і для різних мембран однієї і тієї ж клітини.
Крім ліпідів, мембрани містять також значну частину білків, які плавають у ліпідній двомірній рідині.
- Ліпідний бішар визначає основні структурні особливості біологічних мембран, а типи мембранних білків і їх кількість у мембрані відображають біологічні функції даної мембрани.
Наприклад, мієлінові мембрани нервових клітин виконують функцію електричної ізоляції, тому ці мембрани містять менше 25 % білка. На мембранах мітохондрій та хлоропластів проходить велика кількість ферментативних реакцій, тому ці мембрани містять близько 75 % білків. У звичайній плазматичній мембрані білок становить близько 50 % від її маси.
Залежно від міцності зв’язків між ліпідами і білками, всі мембранні білки можна поділити на дві групи:
· периферійні – ті, що досить легко відділити від мембрани;
· інтегральні – ті, які можна виділити лише після повного розпаду бішару.
Зрозуміло, що мембранні білки, які одночасно контактують як з гідрофільним середовищем всередині чи поза клітиною, так і з гідрофобним середовищем бішару, повинні мати амфіпатичні властивості. Всередині мембран, де вода практично відсутня, полярні групи білків не можуть утворювати водневих зв’язків з молекулами води. Тому вони намагаються утворити їх одна з одною. Ми вже знаємо, що утворення водневих зв’язків вздовж поліпептидного ланцюга призводить до створення a-спіралей чи b-складок. Саме такі форми організації є характерними для мембранних білків.
Білки мембран здатні, як і ліпіди, рухатися у площині бішару (латеральна дифузія). Але швидкість цього руху у десятки і тисячі разів повільніша, ніж у ліпідів. Це пов’язано з тим, що у багатьох клітин є орієнтація типу “верх – низ”. Наприклад, клітини слизових оболонок однією стороною контактують з зовнішнім середовищем, а трьома іншими – з сусідніми клітинами. Зрозуміло, що білковий склад мембран на цих поверхнях має бути різним, тому у мембранах існують спеціальні механізми, які заважають латеральній дифузії білкових молекул у бішарі. Досягти закріплення білків у ліпідній рідині мембран можна трьома різними способами:
· Білкові молекули можуть з’єднуватися одна з одною у великі агрегати. Чим більший такий комплекс, тим повільніше він рухається. Здебільшого швидкість руху може бути майже нульовою.
· Білкові молекули сусідніх клітин можуть з’єднуватися одна з одною.
· Мембранні білки можуть з’єднуватися зі структурами всередині цитозолю.
Враховуючи значну роль білків у біологічних процесах, дослідження особливостей будови та функціонування мембранних білків – один з приоритетних напрямків сучасної біології.
У плазматичній мембрані всіх еукаріотичних клітин більшість білків, розташованих на поверхні клітин, а також більшість ліпідів зовнішнього моношару ковалентно зв’язані з олігосахаридами. Ці вуглеводні ланцюги можуть бути досить складними. Для багатьох різновидностей таких олігосахаридних “хвостів” ще навіть не встановлена точна структура. Функція цих вуглеводів достеменно невідома. Але вважається, що вони відіграють важливу роль у процесах міжклітинного розпізнавання.
Зовнішні вуглеводні компоненти мембран прокаріотичних клітин не лише додатково захищають організми від навколишнього середовища, але і зберігають їх цілісність. Внутрішній тиск цитоплазми у деяких бактерій може перевищувати атмосферний у двадцять і більше разів (1,96 · 106 Па і більше). Наявність додаткової вуглеводної оболонки надає клітинам прокаріотів міцності.
Транспортування речовин через мембрани
Для нормального існування будь-якої складної системи (чи то екосистема, чи людська спільнота) однією з найважливіших проблем є робота транспортних комунікацій. Зі свого життєвого досвіду ви знаєте, що невирішеність цього питання створює значні складності, а іноді навіть може бути причиною катастрофи. Транспортна проблема існує і на рівні клітин. Вона має два аспекти:
· з одного боку, плазматична мембрана у основній своїй масі є гідрофобною, а зовнішнє і внутрішнє середовище і речовини, які транспортуються, є гідрофільними;
· з іншого боку, перенесення речовин досить часто відбувається не лише з місць, де вони знаходяться у високих концентраціях, у місця, де їх концентрація низька, але і навпаки – з низької у високу.
Яким же чином речовини переносяться через плазмолему? Найпростішою відповіддю на це питання були б отвори у мембрані. Але у такому випадку стає неможливим підтримка сталості внутрішнього середовища клітин. Крім того, біліпідний шар взагалі не може мати отворів. Уявіть лише мильну бульбашку з дірочками. Інший гіпотетичний варіант – утворення на поверхні клітини спеціальних ліпосом, які б транспортували речовини у клітину і з клітини. Цей варіант досить прийнятний, але він не може бути універсальним з суто економічних причин. Природа дуже економна господиня, а створення ліпосом і їх переміщення вимагає постійних витрат значної кількості речовини і енергії. У процесі еволюції з’ясувалося, що проблему клітинного транспорту не можливо розв’язати якимось одним методом. Через те, що речовини, необхідні одноклітинним організмам і клітинам багатоклітинних організмів, дуже різні, існує декілька транспортних механізмів (рис. 1.47). Їх можна поділити на дві великі групи:
· транспорт малих молекул;
· транспорт макромолекул і часток.
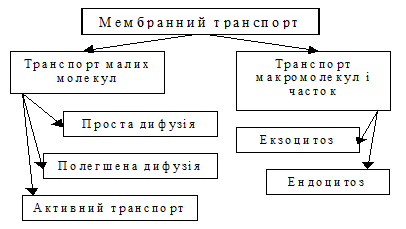
Рис. 3. Види мембранного транспорту
Транспортування малих молекул відбувається на основі закономірностей, які пов’язані з різницею фізико-хімічних характеристик середовища у клітині і за її межами.
- Різницю концентрацій речовин по обидві сторони плазматичної мембрани називають градієнтом концентрації, а різницю електричних зарядів – мембранним потенціалом. Обидва градієнти разом складають електрохімічний градієнт.
У випадку, коли через мембрану проникають неполярні молекули, вони рухаються стосовно концентрації: у напрямку від більшого градієнту до меншого градієнту, до вирівнювання концентрацій. Тому для проникнення у клітину газів (азоту, кисню) та гідрофобних молекул (бензолу) не потрібно ніяких особливих пристосувань.
- Рух хімічних речовин за градієнтом названий пасивним транспортом. Він не потребує додаткових затрат енергії.
- Вид пасивного транспорту, при якому речовини проникають безпосередньо через мембранний ліпідний бішар, називають простою дифузією.
Для полярних молекул ліпідний бішар може бути серйозною перешкодою. Дослідження зі штучними мембранами, які складалися лише з ліпідів, показали, що шляхом простої дифузії можуть проникати лише невеликі незаряджені молекули (вода, сечовина, гліцерол, вуглекислий газ). Коли молекула заряджена (іон), то вона проникає через ліпідний бішар значно важче. Наприклад, коефіцієнт дифузії через біліпідний шар у води у 1010 більший, ніж у іону калію. Великі незаряджені молекули (глюкоза, сахароза) також не проникають через біліпідний шар. Але результати експериментів зі штучними мембранами вступають у протиріччя із реальними фактами. Адже добре відомо, що і глюкоза, і амінокислоти та інші великі полярні молекули й іони можуть проникати через плазматичну мембрану. Без такого транспорту існування у клітині було б неможливе.
Це протиріччя легко спростувати, коли згадати, що у клітинній мембрані, крім ліпідів, є ще і білки. Саме вони виконують роль посередників для транспорту тих молекул, проста дифузія яких через біліпідний шар недостатня для нормального існування клітини.
- Пасивний транспорт, у якому використовуються спеціальні механізми для перенесення речовин, названий полегшеною дифузією.
Білки, які забезпечують полегшену дифузію, можна поділити на два класи:
· до першого класу відносяться білки, що формують канали, через які проникають полярні молекули. Ці білки називають тунельними або каналоутворюючими;
· до другого класу відносяться білки, які зв’язуються з полярними молекулами і переносять їх через мембрану. Їх називають білками-переносниками або транспортерами.
Важливою особливістю пасивного транспорту (простої і полегшеної дифузії) є те, що жоден з цих способів перенесення речовин через клітинну мембрану не потребує затрат енергії. Це пояснюється тим, що рух речовин через мембрану відбувається за градієнтом концентрації та електрохімічним градієнтом. Будь-яке переміщення речовин проти градієнту з більш концентрованих до менш концентрованих, обов’язково вимагає затрат енергії.
- Рух хімічних речовин проти градієнта концентрації чи заряду називається активним транспортом. Він можливий лише при умові використання енергії.
Активний транспорт у клітині відбувається за участю спеціальних, іноді дуже складних, ферментативних комплексів. Як правило, ферменти у таких комплексах зв’язані з мембранами або самі є мембранними білками.
Типовим прикладом спеціального транспортного комплексу є натрій-калієвий насос ((Na+ -K+ )-насос). В усіх тваринних клітинах вміст K+ у цитоплазмі значно перевищує концентрацію цього іону у міжклітинному середовищі. Na+ , навпаки, у значно більших концентраціях знаходиться у міжклітинній рідині, ніж у клітині. Така різниця у концентрації цих іонів має велике значення для функціонування живих організмів. Завдяки (Na+ -K+ )-насосу:
· підтримується мембранний потенціал;
· регулюється об’єм клітин;
· контролюються внутрішньоклітинні концентрації багатьох речовин;
· він має значення для передачі нервового імпульсу;
· він має значення для проникнення у клітину глюкози.
Тому не дивно, що цей вид активного транспорту дуже поширений у клітинах. Було з’ясовано, що на забезпечення роботи (Na+ -K+ )-насоса затрачується близько третини всієї енергії, необхідної тваринній клітині. У нервових клітин ця величина може досягати навіть 70%.
Механізм активного транспорту іонів натрію і калію досить детально вивчений. Дослідження свідчать, що при гідролізі однієї молекули АТФ з клітини викачуються назовні три іони натрію, а в клітину потрапляють два іони калію. Цікавим є той факт, що роль (Na+ -K+ )-насоса в мембранах виконує білок, який є одночасно АТФ-азою (АТФ-аза – це фермент, котрий каталізує гідроліз АТФ). Тому цей білок, який є одночасно і транспортним і каталітичним (ферментом), має назву (Na+ -K+ )-АТФ-аза. Він складається з 2 субодиниць. Каталітичну (ферментативну) активність має субодиниця масою 100000 Да. Друга субодиниця масою 45000 Да – глікопротеїд. Для роботи цього ферменту необхідні іони магнію. Якщо очистити (Na+ -K+ )-АТФ-азу від ліпідів мембрани і перетворити її з мембранного у вільний білок, то вона буде функціонувати лише як фермент, який каталізує розпад АТФ.
Крім (Na+ -K+ )-насоса у плазматичних мембранах присутні й інші види активного транспорту, наприклад, кальцієвий та протонний насоси.
Через свої розміри макромолекули та частки не можуть проникати у клітину за допомогою активного чи пасивного транспорту. У процесі еволюції був вироблений спеціальний механізм, який відповідає за надходження таких великих фрагментів. Для транспорту часток і великих молекул плазматична мембрана утворює невеликих пухирці – везикули (лат. vesicula – “кулька”). У випадку, коли везикули мають досить великі розміри, їх називають вакуолями.
- Процес утворення везикул для транспорту речовий всередину клітини називають ендоцитозом, а назовні – екзоцитозом.
- Ендоцитоз поділяють на фагоцитоз (транспорт твердих речовин) і піноцитоз (транспорт розчинів).
Значний вклад у дослідження ролі ендоцитозу вніс наш співвітчизник І. І. Мечников. Саме він виявив роль цього процесу в знешкодженні інфекційних мікроорганізмів.
Механізми утворення везикул при екзоцитозі і ендоцитозі лише на перший погляд здаються однаковими. Різниця між ними стає очевидною, коли згадати про те, що однією з важливих рис плазматичної мембрани є її асиметричність. При ендоцитозі всередині везикули знаходиться зовнішня (у системі “клітина – навколишнє середовище”) сторона плазматичної мембрани, а при екзоцитозі – навпаки. Завдяки цій різниці обидва види клітинного транспорту можуть регулюватися незалежно один від одного.