Методические указания для студентов специальности «Биоэкология» Печатается в авторской редакции в соответствии с представленным оригинал-макетом
СОДЕРЖАНИЕ: Системная экология (Применение системного анализа в экологии): Методические указания для студентов специальности «Биоэкология» / Сост. Д. И. Башмаков. — Саранск: Изд-во Мордов ун-та, 2004. 32 сУДК 574(076)
Составитель Д. И. Башмаков
Рецензенты:
доктор сельскохозяйственных наук профессор А. В. Каверин;
кандидат биологических наук профессор В, И. Астраданов
Системная экология (Применение системного анализа в экологии): Методические указания для студентов специальности «Биоэкология» / Сост. Д. И. Башмаков. — Саранск: Изд-во Мордов. ун-та, 2004. - 32 с.
Методические указания содержат вопросы становления и развития системных идей в экологии, основы теории систем и системного анализа, приведены элементы системного анализа в экологии и охране окружающей природной среды, рассмотрены основные вещественные, энергетические и информационные процессы в экосистемах, очерчены проблемы моделирования, объяснения и прогнозирования в экологии, описаны основные математические модели популяций, экосистем и биосферы, раскрыта кибернетическая природа, стабильность экосистем, а также надежность природных систем в связи с преобразованием биосферы в техносферу.
Предназначены для студентов IV — V курсов, обучающихся по специальности «Биоэкология», а также всех интересующихся проблемами теоретической экологии.
Печатается по решению научно-методического совета Мордовского государственного университета имени Н. П. Огарева
Учебное издание
Составитель БАШМАКОВ Дмитрий Идрисович
СИСТЕМНАЯ ЭКОЛОГИЯ (ПРИМЕНЕНИЕ СИСТЕМНОГО АНАЛИЗА В ЭКОЛОГИИ)
Методические указания для студентов специальности «Биоэкология»
Печатается в авторской редакции в соответствии с представленным оригинал-макетом
Подписано в печать 12.10.04. Формат 60 х 84 1/16. Бумага офсетная.
Печать офсетная. Гарнитура Тайме. Усл. печ. л. 1,86. Уч.-изд. л. 2,42.
Тираж 250 экз. Заказ № 1835.
Издательство Мордовского университета
Типография Издательства Мордовского университета
430000, г. Саранск, ул. Советская, 24
Введение. Становление и развитие системных идей в экологии
Впечатляющие успехи в развитии науки и техники на какое-то время создали иллюзию полной независимости человека от природы, подвластности всего окружающего человеку. В последнее время на смену таким представлениям приходит общественное осознание кровной заинтересованности человечества во всем, что происходит в окружающем его мире живого, познание необходимости не «покорения природы», а разумной ее эксплуатации. Характерной чертой новой системы взглядов на взаимоотношения человека с природой является понимание возможности необратимых последствий нашей деятельности и вытекающая отсюда острая потребность в прогнозировании, предсказании непосредственных и более отдаленных результатов нашего вмешательства в «естественный порядок вещей».
Существует несколько подходов к предсказанию поведения сложных систем: использование интуиции и богатого опыта исследователя, сравнение с данными экспериментов, проделанных на тождественных или похожих системах, и, наконец, математическое моделирование. В экологических исследованиях должны использоваться — и используются — все возможные подходы. Однако в наше время масштабы и характер вмешательства человека в природные экосистемы столь беспрецедентны, что интуиция исследователя сплошь и рядом отказывает. Возможности же экспериментирования с природными экосистемами по понятным причинам более чем ограниченны. Отсюда ясными становятся важность и актуальность математического моделирования в экологии.
Курс «Системной экологии» имеет целью обобщить полученные студентами за годы обучения в вузе знания по экологии на основе системного подхода к теоретическим вопросам общей экологии и применения системного анализа к решению экологических проблем, дать с единых позиций характеристику всех основных компонентов экосистем, описать их взаимосвязи друг с другом и с внешней средой.
Задача системной экологии состоит в описании принципов, упрощений и абстракций, к которым необходимо научиться сводить многообразие реального мира природы, прежде чем приступить к построению его математических моделей. В этом случае моделирование следует считать специфическим методом системной экологии, с помощью которого исследуются законы функционирования и развития экосистем во времени и пространстве.
Вообще говоря, системный подход не является строго методологической концепцией: он выполняет эвристические функции, ориентируя конкретные экологические исследования в двух основных направлениях. Во-первых, его содержательные принципы позволяют фиксировать недостаточность старых, традиционных методов изучения экосистем для постановки и решения новых задач их целостного исследования. Во-вторых, понятия и принципы конструктивного системного подхода помогают создавать новые программы изучения, ориентированные на раскрытие сущности процессов трансформации энергии, передачи вещества и информации в экосистемах.
Эффективное осуществление методологии системного подхода стало возможным только в середине 60-х гг., когда в распоряжение экологов поступили мощные ЭВМ и были разработаны методы моделирования сложных динамических систем, главным образом в аэрокосмических и технических исследованиях, которые в совокупности получили название системного анализа.
Именно к этому периоду относится появление первых публикаций по математическому моделированию динамики экосистем (биогеоценозов) в нашей стране (Ляпунов, 1966, 1968; Полетаев, 1966; Винберг, Анисимов, 1966; Эман, 1966) и за рубежом (Olson, 1963 - Watt (ed.), 1966; Van Dyne, 1966, 1969; Davidson, Clymer, 1966; Holling, 1966; Levins, 1966; King, Paulik, 1967; Odum, 1967). Начиная с этого времени применение в экологии системного анализа все более расширялось, что сопровождалось усовершенствованием моделей и приемов моделирования, а также обратным влиянием моделирования на стратегию и тактику экологических исследований и даже методологические установки экологов.
Успехи в изучении и моделировании экосистем, способствовали окончательному утверждению системной парадигмы, опирающейся на концепцию экосистемы.
? Вопросы к семинару ?
1) Каковы цели и задачи системной экологии? Что можно принять за метод системной экологии?
2) Что такое системный подход и когда он появился?
3) История формирования системных идей в экологии.
Тема 1. Основы теории систем и системного анализа Системы и закономерности их формирования и развития
1. Система. Простые и сложные системы. Классификация систем
«Система» (от греч. systema - целое, составленное из частей) считается одним из ключевых философско-методологических и специальных научных понятий. Система -совокупность элементов со связями между ними. Элемент системы из-за иерархической структуры мира сам оказывается системой со своими элементами. Фиксация системы делит мир на две части - на систему и среду. При этом подчеркивается большая сила связей элементов внутри системы по сравнению с силой связей с элементами среды.
Для идентификации систем необходимо задать еще один критерий, который Ю.А. Урманцев назвал «законом композиции». Выбор закона композиции даст возможность объединить те же объекты в разные системы.
Каждая система определяется некоторой структурой (элементы и взаимосвязи между ними) и поведением (изменение системы во времени).
Сложность системы на «структурном уровне» задается числом ее элементов и связей между ними. Дать определение «сложности» в этом случае чрезвычайно трудно: исследователь сталкивается с так называемым «эффектом кучи» (один шар - не куча, два шара - не куча, три - не куча, а вот сто шаров - куча, девяносто девять - куча; так где же граница между «кучей» и «не кучей»?).
Определить, что такое «сложная система на «поведенческом уровне» представляется более реалистичным.
Б.С. Флейшман (1978, 1982) предложил пять принципов усложняюиегося поведения систем, представленных на рис. 1.
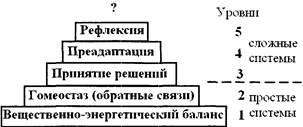
Рис. 1. Принципы усложнения систем на поведенческом уровне
Системы, включающие в себя в качестве хотя бы одной подсистемы решающую систему (поведению которой присущ акт решения), называют сложными (системы 3-5 уровней; такие системы изучает системология). Классификация систем представлена на рисунке 2.
2. Иерархия уровней организации
Иерархия — это «расположение ступенчатым рядом». На каждой ступени, или уровне, в результате взаимодействия с окружающей физической средой (энергией и веществом) возникают характерные функциональные системы.
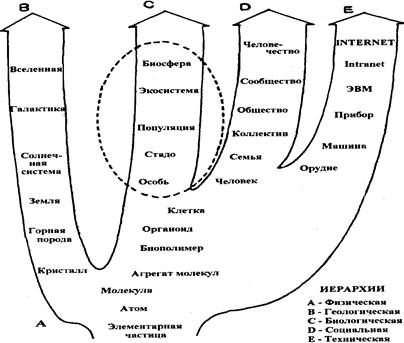
Рис. 2, Иерархии природных систем
Под системой подразумевают также «упорядоченные взаимодействующие и взаимозависимые компоненты, образующие единое целое»; в соответствии с другой точкой зрения система - это «набор взаимосвязей, составляющий поддающуюся идентификации единицу, реальную или концептуальную». Системы, содержащие живые компоненты ( биологические системы или биосистемы) показаны на рисунок 3.
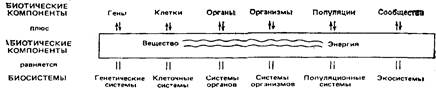
Рис. 3. Иерархия биосистем
Экология изучает системы, расположенные в правой части этого спектра, т.е. системы выше уровня организма. В экологии значение термина популяция, первоначально обозначавшего группу людей, расширено и обозначает группы особей любого вида организмов. Сообщество включает все популяции, занимающие данный участок. Сообщество и неживая среда функционируют совместно, образуя экологическую систему, или экосистему. Сообществу и экосистеме приблизительно соответствуют часто употребляемые в европейской и русской литературе термины биоценоз и биогеоценоз (буквально
жизнь и земля, функционирующие вместе). Биом — крупная региональная или субконтинентальная биосистема, характеризующаяся каким-либо основным типом растительности или другой характерной особенностью ландшафта. Самая крупная и наиболее близкая к идеалу в смысле «самообеспечения» биологическая система, которую мы знаем, — это биосфера, или экосфера; она включает все живые организмы Земли, находящиеся во взаимодействии с физической средой Земли как единое целое, чтобы поддерживать эту систему в состоянии устойчивого равновесия, получая поток энергии от Солнца, ее источника, и переизлучая эту энергию в космическое пространство. Под устойчивым равновесием мы понимаем способность саморегулируемой системы возвращаться в исходное состояние по крайней мере после небольшого отклонения.
3. Иерархический, сетевой и реляционный подходы к анализу систем
На самых верхних и самых нижних этажах экологической иерархии иерархический принцип классификации работает неплохо. Традиционно выделяемые в экологии группы продуцентов, редуцентов и консументов (экоцарства) достаточно легко и естественно делятся на экотипы.
С другой стороны, выделив эковиды как совокупности экологически однородных особей, занимающих элементарные фундаментальные ниши, мы можем достаточно уверенно объединить их в экороды. Ведь для выполнения одной и той же роли в реальных экосистемах имеется, как правило, несколько эковидов, различающихся оптимальными условиями своего существования.
Но на уровне экосемейств, экоотрядов и экоклассов возможно построение нескольких иерархий, в основе которых лежат различные экологические признаки. Поэтому внутри отдельных экотипов возможно построение самых разных иерархий, сам иерархический подход на этих уровнях теряет свою руководящую роль, уступая место подходу сетевому. Если иерархическая структура древовидна, то в сетевых структурах любой элемент может быть в принципе связан с любым другим элементом.
Однако, классификацию, построенную по сетевому принципу, можно эффективно применять на практике лишь с помощью компьютерной техники. Реляционный подход позволяет свести всю переработку информации, заложенной в классификационной схеме, к манипуляциям с матрицами. Каждая двумерная таблица, называемая реляцией или отношением, наглядно представляет наложение любых двух иерархий, входящих в сетевую структуру, на определенном их уровне, что позволяет последовательно просматривать все возможные варианты.
4. Основные принципы системологии
Принцип эмерджентности. По мере объединения компонентов, или подмножеств, в более крупные функциональные единицы, у этих новых единиц возникают новые свойства, отсутствовавшие на предыдущем уровне. Эмерджентные свойства экологической единицы нельзя предсказать, исходя из «свойств компонентов, составляющих эту единицу. При каждом объединении подмножеств в новое множество возникает по меньшей мере одно новое свойство.
Принцип иерархической организации (или принцип интегративных уровней Одума): позволяет соподчинить друг другу как естественные, так и искусственные системы.
Принцип несовместимости Л. Заде : сложность системы и точность, с которой ее можно анализировать, связаны обратной зависимостью.
Принцип контринтуитив ного поведения Дж. Форрестера : дать удовлетворительный прогноз поведения сложной системы на достаточно большом промежутке времени, опираясь только на собственный опыт и интуицию практически невозможно.
Принцип множественности моделей В.В. Налимова : для объяснения и предсказания структуры и (или) поведения сложной системы возможно построение нескольких моделей, имеющих одинаковое право на существование.
Принцип осуществимости Б.С. Флейшмана : мы не надеемся на везение и у нас мало времени. Системология рассматривает только те модели, для которых алгоритм осуществим, т.е. решение может быть найдено с заданной вероятностью р0 время to.
Принцип Формирования законов : законы системологии носят дедуктивный характер и никакие реальные явления не могут опровергнуть или подтвердить их справедливость.
Принцип рекуррентного объяснения свойства систем данного уровня иерархической организации мира объясняются, исходя из постулируемых свойств элементов этой системы и связей между ними.
Принцип минимаксного построения моделей : теория должна состоять из простых моделей (min) систем нарастающей сложности (max).
? Вопросы к семинару ?
1) Что такое «система»? Что такое «сложная система»?
2) Иерархия уровней организации природных систем.
3) Иерархический, сетевой и реляционный подходы к анализу систем, их особенности и область применения.
4) Основные принципы системологии: принцип эмерджентности, принцип иерархической организации, принцип несовместимости, принцип контринтуитивного поведения, принцип множественности моделей, принцип осуществимости, принцип формирования законов, принцип рекуррентного объяснения, принцип минимаксного построения моделей и др.
Тема 2. Элементы системного анализа в экологии и охране окружающей природной среды
Концепция экосистем по Ю. Одуму является главенствующей в современной экологии. Любая биосистема, включающая все совместно функционирующие организмы на данном участке и взаимодействующая с физической средой таким образом, что поток энергии создает четко определенные биотические структуры и круговорот веществ между живой в неживой частями, представляет собой экосистему. Экосистема — основная функциональная единица в экологии. 1. Структура экосистемы
С точки зрения трофической структуры (от греч. trophe — питание) экосистему можно разделить на два яруса: 1) верхний автотрофный (самостоятельно питающийся) ярус, или «зеленый пояс», включающий растения или их части, содержащие хлорофилл, где преобладают фиксация энергии света, использование простых неорганических соединений и накопление сложных органических соединений, и 2) нижний гетеротрофный (питаемый другими) ярус, или «коричневый пояс» почв и осадков, разлагающихся веществ, корней и т. д., в котором преобладают использование, трансформация и разложение сложных соединений. С биологической точки зрения в составе экосистемы удобно выделять следующие компоненты: 1) неорганические вещества (С, N, С02 , Н2 0 и др.), включающиеся в круговороты; 2) органические соединения (белки, углеводы, липиды, гумусовые вещества и т. д.), связывающие биотическую и абиотическую части; 3) воздушную, водную и субстратную среду, включающую климатический режим и другие физические факторы; 4) продуцентов, автотрофных организмов, в основном зеленые растения, которые могут производить пищу из простых неорганических веществ; 5) макроконсументов, или фаготрофов (от греч. phagos— пожиратель),— гетеротрофных организмов, в основном животных, питающихся другими организмами или частицами органического вещества; 6) микроконсументов, сапро-трофов (от греч. sapros — гнилой), деструкторов, или осмотрофов (от греч. osmos— толчок, давление),— гетеротрофных организмов, в основном бактерий и грибов, получающих энергию либо путем разложения мертвых тканей, либо путем поглощения растворенного органического вещества, выделяющегося самопроизвольно или извлеченного сапротрофами из растений и других организмов.
Для функционирования экосистемы и поддержания ее структуры необходимо взаимодействие трех основных компонентов, а именно сообщества, потока энергии и
круговорота веществ.
2. Гипотеза однонаправленности потока энергии
ГИПОТЕЗА ОДНОНАПРАВЛЕННОСТИ ПОТОКА ЭНЕРГИИ - представление о потоке энергии через продуценты к консументам и редуцентам с падением величины потока на каждом трофическом уровне (в результате процессов жизнедеятельности). Часть поступающей солнечной энергии преобразуется сообществом и переходит на качественно более высокую ступень, трансформируясь в органическое вещество, представляющее собой более концентрированную форму энергии, чем солнечный свет, но большая часть энергии деградирует, проходит через систему и покидает ее в виде низкокачественной тепловой энергии (тепловой сток).
Материя циркулирует в системе, а энергия — нет. Живые замкнутые термодинамические системы невозможны. Все экосистемы, даже самая крупная — биосфера, являются открытыми системами: они должны получать и отдавать энергию. Поэтому в концептуально законченную экосистему входит среда на входе и среда на выходе и система, т. е. Экосистема = IE + S + OE .
? Вопросы к семинару ?
1) Экосистема и биогеоценоз. Основные сходства и различия этих понятий.
2) Основные виды и формы представления структур экосистем.
3) Гипотеза однонаправленности потока энергии.
Тема 3. Экология биосферы. Вещественные, энергетические и информационные процессы в экосистемах
Связь компонентов экосистемы осуществляется через три качественно разных группы процессов - преобразование и перемещение вещества, расходование энергии и ее накопление в связанной форме. Упорядочение этих процессов может рассматриваться как передача, накопление и перекодирование информации.
1. Типы земного вещества
В.И. Вернадский (1926) выделил 7 типов вещества биосферы: живое вещество, биогенное вещество (каустобиолиты, известняки и т.д.), косное вещество (образуется процессами, в которых живые организмы не участвуют), биокосное вещество (создается одновременно живыми организмами и процессами неживой природы), радиоактивное вещество, рассеянные атомы и вещество космического происхождения.
Центральное звено в концепции В.И. Вернадского о биосфере - представление о живом веществе. Живые организмы являются функцией биосферы и теснейшим образом материально и энергетически с ней связаны, являются огромной геологической силой, ее определяющей. Количество энергии, заключенное в живом веществе равно 4,191021 -4,19-1022 Дж (1018 -1019 ккал). Значительная часть ее идет на образование в пределах биосферы новых минералов, вне биосферы неизвестных (граниты), а часть -захоранивается в виде самого органического вещества (залежи каустобиолитов, нефти, газа). То есть при участии живого вещества лучистая энергия Солнца медленно проникает в глубь планеты. Благодаря деятельности живых организмов образуются кора выветривания и почва, формируется определенный химический состав подземных и поверхностных вод, поддерживается баланс газов в атмосфере.
Живое вещество распределено в биосфере крайне неравномерно. В зависимости от занимаемой площади В.И. Вернадский различал жизненные пленки (прослеживаются на огромных расстояниях - планктонные сообщества поверхности океана) и сгущения жизни (более локальные скопления - например, Саргассово море).
2. Основные экологические законы, связанные с веществом биосферы ПРАВИЛА БЕЙЕРИНКА (Bejerinck): а) все есть всюду (бактерии - жизненная
пленка - развиваются повсюду, где есть условия для их существования; и б) среда отбирает (организмы отобраны средой либо потому, что в данных условиях могут развиваться только эти организмы, либо потому, что они побеждают своих конкурентов).
ГИПОТЕЗА КОНСТАНТНОСТИ Вернадского - количество живого вещества биосферы для данного геологического периода есть величина постоянная.
АКСИОМА БИОГЕННОЙ МИГРАЦИИ АТОМОВ Вернадского - миграция химических элементов на земной поверхности и в биосфере осуществляется или непосредственно при участии живого вещества, или протекает в среде, геохимические свойства которой обусловлены живым веществом.
ПРОПОРЦИЯ (УРАВНЕНИЕ) РЭДФИЛДА - соотношение атомов важнейших элементов в биосфере имеет следующий вид: C:N:P = 100:15:1.
БИОГЕОХИМИЧЕСКИЕ ПРИНЦИПЫ Вернадского:
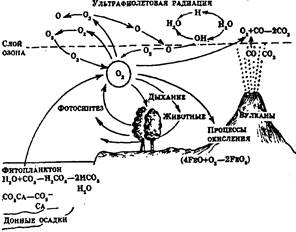
Рис. 4. Круговорот кислорода в природе
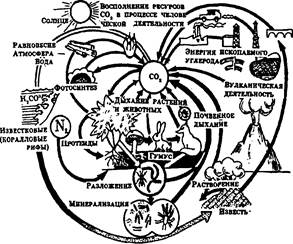
Рис. 5. Круговорот углерода в природе
1) Биогенная миграция атомов химических элементов в биосфере всегда стремится к максимальному своему проявлению.
2) Эволюция видов в ходе геологического времени, приводящая к созданию форм жизни, устойчивых в биосфере, идет в направлении, увеличивающем биогенную миграцию атомов биосферы.
3) В течение всего геологического времени, с криптозоя (докембрий), заселение планеты должно было быть максимально возможным для всего живого вещества, которое тогда существовало.
3. Главные биогеохимические циклы биосферы
Самая существенная особенность биосферы -биогенная миграция атомов химических элементов (см. аксиому биогенной миграции атомов). Под биологическим круговоротом понимается поступление химических элементов их почвы и атмосферы в живые организмы; превращение в них поступающих элементов в новые сложные соединения и возвращение их в почву и атмосферу и в воду в процессе жизнедеятельности с ежегодным спадом части органиче-
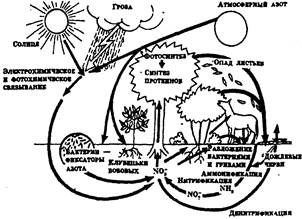
Рис. 6. Круговорот азота в природе
ского вещества или с полностью отмершими организмами, входящими в состав биогеоценоза.
Биогеохимический круговорот и биогеохимические связи суши, моря, атмосферы, почвы, пресных вод и организмов весьма сложны. Каждый элемент или вещество имеет свою собственную структуру биогеохимического круговорота, отличающегося, по крайней мере в количественных деталях, от циркуляции всех других элементов.
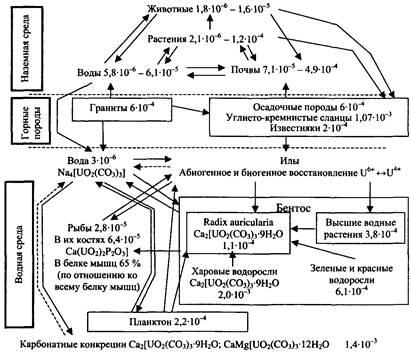
Рис. 7. Круговорот урана (значения даны в %)
Все эти циклы, дополняемые циклами воздуха и воды, которые являются важной составной частью механизмов круговорота веществ, дают основание говорить о том, что локальные экосистемы земного шара образуют вместе единую мировую экосистему -биосферу. Имитационные модели основных биогеохимических циклов представлены на рисунках 4-7.
? Вопросы к семинару ?
1) Основные типы земного вещества и их характеристика. Живое вещество как ведущая сила эволюции земной природы.
2) Основные экологические законы, связанные с веществом биосферы: правила Бейеринка, гипотеза константности, биогеохимические принципы В.И. Вернадского, аксиома биогенной миграции атомов, уравнение Рэдфилда.
3) Система биомов, Гипотеза абиссальных сгущений жизни.
4) Круговороты вещества: большой и малый. Их особенности, скорости и функции. Особенности круговоротов загрязняющих веществ.
5) Круговороты воды и кислорода.
6) Круговороты углерода и азота.
7) Круговороты фосфора, серы и урана.
4. Термодинамика экосистем: закон энтропии
Экология изучает связь между светом и экологическими системами и способы превращения энергии внутри системы. Энергию определяют как способность производить работу. Свойства энергии описываются следующими законами.
Первый закон термодинамики, или закон сохранения энергии, гласит, что энергия может переходить из одной формы в другую, но она не исчезает и не создается заново.
Второй закон термодинамики, или закон энтропии, формулируется по-разному, в частности таким образом: поскольку некоторая часть энергии всегда рассеивается в виде недоступной для использования тепловой энергии, эффективность самопроизвольного превращения кинетической энергии (например, света) в потенциальную (например, энергию химических соединений протоплазмы) всегда меньше 100 %. Мера количества связанной энергии, которая становится недоступной для использования -энтропия (от греч. entropia - поворот, превращение). Этот термин также используется как мера изменения упорядоченности, которая происходит при деградации энергии.
Важнейшая термодинамическая характеристика организмов, экосистем и биосферы в целом - способность создавать и поддерживать высокую степень внутренней упорядоченности, т.е. состояние с низкой энтропией. И. Пригожин (1962) показал, что способность к самоорганизации и созданию новых структур встречается в системах, далеких от равновесия и обладающих хорошо развитыми «диссипативными структурами», откачивающими неупорядоченность.
Экосистемы и организмы представляют собой открытые неравновесные термодинамические системы, постоянно обменивающиеся с окружающей средой энергией и веществом, уменьшая этим энтропию внутри себя, но увеличивая энтропию вовне в согласии с законами термодинамики.
Термин «энтропия» используется и в более широком смысле - для обозначения деградации различных материалов. Так, недавно выплавленная сталь - это низкоэнтропийное состояние железа, а ржавеющий кузов автомобиля - высокоэнтропийное. Соответственно для «высокоэнтропийного» человеческого общества характерна деградация энергии, ржавеющая техника, лопающиеся водопроводные трубы и разрушаемая эрозией почва. Постоянные восстановительные работы - неизбежная плата за цивилизацию с высоким расходом энергии.
5. Экологические законы, связанные с энергетическими потоками биосферы ПРИНЦИП ЛЕ ШАТАЛЬЕ-БРАУНА - при внешнем воздействии, выводящем
систему из состояния устойчивого равновесия, равновесие смещается в том направлении, в котором эффект внешнего воздействия ослабляется.
Следствием из принципа Ле Шаталъе - Брауна) является ЗАКОН ТОРМОЖЕНИЯ РАЗВИТИЯ - в период наибольших потенциальных темпов развития системы возникают максимальные тормозящие эффекты.
Принцип Ле Шаталье-Брауна применим в рамках классической физики для описания процессов в закрытых системах (не получающих энергии извне); так как экосистемы — принципиально открытые системы (обмениваются энергией, веществом, информацией с окружающей средой), то для их описания более корректными выглядят представления теории нелинейных необратимых процессов. Для закрытых систем общим принципом является второе начало термодинамики, для открытых - ПРИНЦИП НЕРАВНОВЕСНОЙ ДИНАМИКИ ПРИГОЖИНА-ОНСАГЕРА - Неравновесность есть то, что порождает «порядок из хаоса». Если закрытые системы имеют одно состояние равновесия, то открытые - несколько. Перейдя границу устойчивости система попадает в критическое состояние, называемое точкой бифуркации. В этой точке даже небольшая флуктуация может вывести систему на иной путь эволюции и резко изменить ее структуру и поведение.
ПОСТУЛАТ МАКСИМУМА БИОГЕННОЙ ЭНЕРГИИ Вернадского-Бауэра - любая экосистема, находясь в состоянии устойчивого неравновесия (т.е. динамического подвижного равновесия с окружающей средой) и эволюционно развиваясь, увеличивает свое воздействие на среду.
ЗАКОН ПИРАМИДЫ ЧИСЕЛ Элтона (1927) - число индивидуумов в последовательности трофических уровней убывает и формирует пирамиду чисел.
ЗАКОНПИРАМИДЫ БИОМАСС (Одум, 1975). Пирамиды биомасс представляют более фундаментальный интерес, так как они дают ...картину общего влияния отношений в пищевой цепи на экологическую группу как целое.
ЗАКОН ПИРАМИДЫ ПРОДУКТИВНОСТИ - более стабильная пирамида, чем пирамида чисел или пирамида биомасс, которая в значительно большей степени отражает последовательность трофических уровней.
ПРАВИЛО ДЕСЯТИ ПРОЦЕНТОВ (пирамида энергий Станчинского) - средне-максимальный переход 10 % энергии (или вещества в энергетическом выражении) с одного трофического уровня экологической пирамиды на другой, как правило, не ведет к неблагоприятным для экосистемы в целом и теряющего энергию трофического уровня последствиям.
АКСИОМА ЭКОЛОГИЧЕСКОЙ АККУМУЛЯЦИИ ЭНЕРГИИ - часть проходящей через экосистему энергии накапливается и временно выключается из общего энергетического потока.
ПРАВИЛО ОДНОГО ПРОЦЕНТА В.Г. Горшкова (1985) - изменение энергетики природной системы на 1%, как правило, выводит природную систему из равновесного (квазистационарного) состояния.
ПРИНЦИП МАКСИМИЗАЦИИ ЭНЕРГИИ Лотки-Одума-Пинкертона - в «соперничестве» с другими экологическими объектами выживают (сохраняются) те из них, которые наилучшим образом способствуют поступлению энергии и используют максимальное ее количество наиболее эффективным способом. 6. Элементы биоэнергетики экосистем
Особая роль растительности в общей структуре живой природы связана с основной функцией растительного покрова нашей планеты - аккумуляцией и превращением солнечной энергии в энергию химических связей органического вещества с последующей передачей ее тем компонентам экосистемы, которые не в состоянии самостоятельно фиксировать энергию Солнца.
Установлено, что к верхним слоям атмосферы Земли от Солнца приходят 1,94 кал/см2 в минуту, из которых биосферы достигает только около 0,9 кал/см2 /мин, а поверхности Земли - менее 0,3 кал/скг/мин. В средних широтах каждый гектар поверхности планеты получает 9-10 кал/год. Однако верхний предел фиксации солнечной энергии растительность составляет всего 5 % от посылаемой Солнцем энергии.
Энергия в экосистеме на примере смешанного леса представлена на рисунке 8.
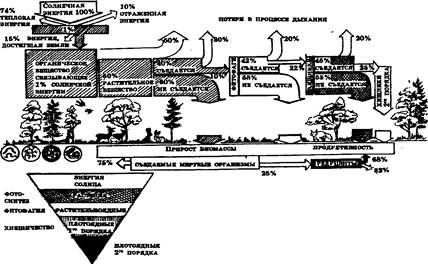
Рис. 8. Энергия в экосистеме на примере смешанного леса
Всякий источник энергии, уменьшающий затраты на самоподдержание экосистемы и увеличивающий ту долю энергии, которая может перейти в продукцию, называется вспомогательным потоком энергии, или энергетической субсидией.
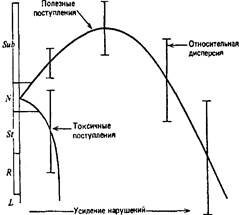
Фактор, который при одних условиях среды или при одном уровне поступлений увеличивает продуктивность, при других условиях среды или другом уровне поступлений может способствовать утечкам энергии, уменьшая продуктивность (рисунок 9).
Обобщенная кривая, показывающая, как увеличенное поступление энергии или веще-
Рис. 9. Кривые субсидии и стресса ства может вывести систему из
диапазона обычного функционирования ( N ). Если система может использовать этот излишек, то ее основные функции, например продуктивность, при умеренных уровнях повышения притока могут усилиться (эффект субсидии— Sub ), но при дальнейшем увеличении притока эти функции начинают подавляться (эффект стресса — St ). Если поступают ядовитые вещества, функции подавляются и высока вероятность того, что сообщество заменится другим, более толерантным, или экосистема вообще погибнет. R — замещение, L — гибель.
7. Концепция продуктивности
Одна из наиболее важных характеристик биоэнергетики фитоценозов - скорость фиксации и накопления органического вещества, характеризующая продуктивность растительных сообществ.
Первичная продуктивность экологической системы, сообщества или любой их части определяется как скорость, с которой лучистая энергия усваивается организмами-продуцентами в процессе фото- и хемосинтеза, накапливаясь в форме органических веществ. В процессе производства органического вещества выделяют четыре последовательных уровня:
1. Валовая первичная продуктивность (валовый фотосинтез, общая ассимиляция) — это общая скорость фотосинтеза, включая те органические вещества, которые за время измерений были израсходованы на дыхание.
2. Чистая первичная продуктивность (наблюдаемый фотосинтез, чистая ассимиляция) — скорость накопления органического вещества в растительных тканях за вычетом того органического вещества, которое использовалось при дыхании растений за изучаемый период.
3. Чистая продуктивность сообщества — скорость накопления органического вещества, не потребленного гетеротрофами за учетный период (за вегетационный период, год).
4. Вторичная продуктивность - скорость накопления энергии на уровнях консу-ментов.
? Вопросы к семинару ?
1) Законы термодинамики в применение к природным (биосистемам, антропогенным) системам.
2) Свойства закрытых и открытых систем. Принцип неравновесной термодинамики Пригожина-Онсагера.
3) Элементы биоэнергетики экосистем. Концепция продуктивности.
4) Основные экологические законы, связанные с энергетическими потоками биосферы: постулат максимума биогенной энергии, законы экологических пирамид, правило десяти процентов, правило одного процента, принцип максимизации энергии.
5) Концепция энергетической субсидии.
8. Концепция информации
Понятие «информация» невозможно вывести из представлений, относящихся к миру вещей, информация - не материя и не энергия. Безуспешными оказались и попытки рассматривать информацию как третью ипостась материи, наряду с массой и энергией: такая точка зрения неизбежно приводит к выводу о «всюдности» информации, весьма быстро приобретающему мистическую окраску. Поэтому определение информации через описание форм ее проявления и ее свойств представл»лея вполне правомочным. Формой проявления информации является оператор - необходимый компонент целенаправленного действия. Информация - совокупность приемов, правил или сведений, необходимых для построения оператора.
Прием, создание, хранение, передачу и использование информации называют элементарными информационными актами, а осуществление всей совокупности таких актов - информационным процессом. Совокупность механизмов, обеспечивающих полное осуществление информационного процесса, называют информационной системой. Формально информационные системы можно подразделить на закрытые, способные только создавать, хранить и использовать свою информацию, и открытые, способные также ее воспринимать и передавать. Вне информационной системы информация может лишь сохраняться в виде записей на тех или иных физических, носителях, но не может быть ни принятой, ни переданной, ни использованной.
Первой ставшей нам известной информационной системой был человек. Специфическую для человека информацию, которой обмениваются люди при помощи устной
и письменной речи, обычно называют знанием (логическая информация).
Информацией называют также те сведения, которыми обмениваются между собой животные и которые, будучи восприняты, существенно влияют на их поведение. Это - поведенческая информация. Информационными системами, оперирующими с такой информацией, являются все многоклеточные животные, включая человека, и некоторые одноклеточные.
Третий известный нам вид информации - генетическая - записана в нуклеиновых кислотах клеток последовательностью оснований и определяет фенотипические особенности всех без исключения живых существ. Специфическими для этой информации системами являются все негенетические компоненты живых организмов.
Можно утверждать, что вне живой природы информации не существует.
Понятие «информация» несет в себе два оттенка. Во-первых, количественный аспект, игнорирующий смысл и значение сообщения. Предполагается, что различия сообщений связаны только с расположением символов в них и это расположение должно сохраниться при передаче информации. Этим аспектом занимается теория информации, основоположниками которой являются В.А. Котельников и К. Шеннон. Во-вторых, это смысловая оценка сообщения или, как его иногда называют семантический аспект информации (развитие этого направления связано с именами Р. Карнапа и И. Бар-Хиллела).
Свойства информации:
- Фиксируемость. Информация может существовать только в зафиксированном состоянии. Можно полагать, что способов записи информации может существовать столько, сколько может быть способов ее считывания. Для фиксации информации можно использовать не менее двух различных знаков или букв.
- Инвариантность информации означает, что одна и та же информация, независимо от ее семантики, может быть «записана» на любом языке, любым алфавитом, т.е. системой знаков, наносимых любыми способами на любые носители.
- Бренность информации. Поскольку информация всегда зафиксирована на каком-либо физическом носителе, постольку сохранность и само существование информации целиком и полностью определяется судьбой ее носителя. Условием неограниченно-длительного существования информации из-за её бренности является только периодическая ее репликация или, точнее, требование, чтобы скорость репликации была больше скорости ее деградации.
- Транслируемость - возможность быть переданной с одного носителя на другой, такой же или иной физической природы, в той же или иной системе записи.
- Когда скорость транслируемое™ превосходит скорость разрушения и гибели информации, это приводит к ее размножаемости . Следствием размножения информации является ее мультипликативность , т.е. возможность одновременного существования одной и той же информации в виде некоторого числа идентичных копий на одинаковых или разных носителях.
- Изменяемость. Деформируемость физических носителей, а также ошибки при трансляции могут приводить не только к гибели информации, но и к ее изменениям.
- Действенность информации может выявляться лишь в адекватной ей информационной системе, - вне таковой любая информация абсолютно пассивна.
- Семантика информации может проявляться лишь одним путем - в специфике кодируемого ею оператора. Возможность быть использованной - в ее материализованном виде, т.е. в качестве оператора, - для достижения той или иной цели обусловливает
ценность информации.
- Полипотентность - любой оператор может быть использован для достижения разных целей. Из свойства полипотентности следует, что для чего-нибудь полезной может оказаться любая информация.
9. Информация в растительных сообществах
Определение количества информации, которое передается и воспринимается объектами растительного сообщества, еще не дает полного представления о характере взаимодействия этих объектов. Действительно, два воздействия АВ и ВА несут одина-
ковое количество информации, но могут быть причиной различного поведения системы, находящейся под воздействием. Важное значение имеет не только количественная оценка информации, но и ее качественная составляющая (в данном случае - порядок воздействия).
Возможные носители информации в растительных сообществах: 1) качестве информационных сигналов могут выступать различные лимитирующие факторы, 2) факторы, зависящие от плотности особей, 3) аллелохимические (аллелопатические) взаимодействия, 4) электромагнитные поля биологического происхождения.
10. Информационные поля животных
Информационные взаимодействия в животном мире, как и вещественно-энергетические, наиболее отчетливо проявляются на уровне популяции, как совокупность внутри- и межпопуляционных информационных потоков. На популяционном же уровне процессы информации тесно связаны с динамикой плотности населения.
Количественная сторона информации сообщает животным о плотности населения. Каждая особь является приемником информации и ее продуцентом. Эта информация и вызывает ответную реакцию, направленную на восстановление оптимального уровня интенсивности сигнального биологического поля, а, следовательно, и оптимального уровня численности.
По Н.П. Наумову, сигнальное биологическое поле - это упорядоченная совокупность изменения организмами исходных физических и химических характеристик мест их обитания, выступающих как совокупность сигналов или система связей, сплачивающих животных в более или менее тесные группы с согласованным поведением. При конкретных экологических исследованиях сигнальное биологическое поле удобно рассматривать как информационную систему, имеющую три параметра:
- величина биологического поля выступает как нечто, организованное из элементов объектов и событий среды, вовлеченных животным в процессе двигательной активности в сферу своей деятельности. Это пространственный параметр биологического сигнального поля;
- элементами «напряженности биологического поля» выступают уже не физические объекты и события среды, имеющие сигнальное значение для реагирующих на них особей, а реакции особи на них.
- параметр «анизотропность биологического поля» подразумевает субсистему, организованную одновременно из элементов физико-химической природы (объектов и событий среды) и функциональных элементов (повторных реагирований на объекты и события среды сходной сигнальной природы, т.е. несущих одинаковый для воспринимающей его особи сигнал).
Сигнальные биологические поля имеют иерархическую организацию соответствующую системной организации живого надорганизменных уровней: индивидуальное (или элементарное) биологическое поле; групповое биологическое поле; общее сигнальное поле популяции; сигнальное поле коадаптивного комплекса экологически близких видов; биоценотическое сигнальное поле.
11. Информация и феномен жизни
Принципиальное отличие живых объектов от неживых состоит в той, что все живые объекты способны осуществлять целенаправленные действия. Способность эта обусловлена их организацией, особенности которой задаются кодирующей эту организацию информацией. Все живые объекты, по существу, - это информационные системы, которые, попадая в подходящие условия, могут обеспечивать воспроизведение кодирующей их информации. Жизнь, таким образом, - это форма существования информации и кодируемых ею операторов, обеспечивающих возможность воспроизведения этой информации в подходящих для этого условиях внешней среды. Цель жизнедеятельности всех живых организмов - это воспроизведение кодирующей их информации.
В ходе эволюции жизни на Земле возникали информационные системы все большей степени сложности. Феномен жизни и ее эволюцию можно интерпретировать как строго преемственный процесс возникновения и развития информации, постепенно,
по мере исчерпания емкости своих физических носителей, приобретавшей все новые формы: генетической, поведенческой и логической.
? Вопросы к семинару ?
1) Понятие информации, классификация информации, аспекты информации.
2) Основные свойства информации.
3) Информация и феномен жизни. Отличия живого от неживого, отличия человека от других живых организмов.
4) Информация в растительных сообществах.
5) Информационные поля животных.
Тема 4. Модели и моделирование в экологии
1. Модели
По мере развития науки и техники Человек все чаще сталкивается с необходимостью исследования объектов, прямое экспериментирование с которыми невозможно. В подобных ситуациях математическое моделирование и экспериментирование с системами математических моделей, которые с определенной точностью воспроизводят (имитируют) реальность, становятся единственным возможным средством анализа.
Модель - упрощенное, «упакованное» знание, несущее вполне определенную и ограниченную информацию о том или ином предмете, явлении, отражающее те или иные его отдельные свойства. Это упрощение (огрубление) осуществляется путем сознательного удаления из системы ^некоторых элементов и связей, в результате мы получаем подсистему Y1 . С другой стороны, модель должна, в определенном смысле, верно отражать оригинал.
Стратегия моделирования заключается в попытке путем упрощения получить модель, свойства и поведение которой можно было бы эффективно изучать, но которая в то же время оставалась бы достаточно сходной с оригиналом, чтобы результаты этого изучения все же были применимы и к оригиналу. Обратный переход от модели Y к оригиналу называется интерпретацией модели.
Модель, какой бы язык она ни использовала, содержит не только ту информацию, которая послужила ее источником и основой, — в модели оказывается закодированными и новые знания, то, что люди раньше и не знали.
Цели построения моделей: 1) для определения общего направления исследований или для того, чтобы предварительно обрисовать контуры проблемы, подлежащие более детальному изучению; 2) для предсказания изменения системы во времени и в пространстве.
Модели можно оценивать по нескольким основным свойствам:
1) Реалистичность - это степень, с которой математические утверждения модели, будучи облечены в слова, соответствуют биологическим представлениям, которые они призваны отражать.
2) Точность - способность модели количественно предсказывать изменения и имитировать данные, на которых они основаны.
3) Общность - это диапазон приложимости модели, то есть число различных ситуаций, в которых модель может работать.
4) Разрешающая способность - количество признаков системы, которые пытается отразить модель.
2. Анатомия математических моделей
Следует выделять 4 основных компонента математических моделей:
1) системные переменные - это ряды чисел, которые используются для представления состояния системы в любой момент времени. В любой момент времени экосистемы состоят из ряда компонентов (или блоков), для характеристики которых используется одна или несколько системных переменных;
2) взаимоотношение (взаимодействие) между блоками описываются при помощи
функциональных зависимостей, обычно это либо математические формулы, либо уравнения;
3) входы системы (или факторы), которые влияют на компоненты экосистемы, но не находятся под их влиянием описываются вынуждающими функциями;
4) константы математических моделей называются параметрами. Как параметры можно рассматривать и мало изменяющиеся переменные состояния. В свою очередь, параметры могут переходить в переменные, если они не удовлетворяют модели.
3. Классификация моделей
В зависимости от особенностей системы-оригинала и задач исследования применяются самые разнообразные модели, которые целесообразно классифицировать по следующим признакам (рисунок 10).
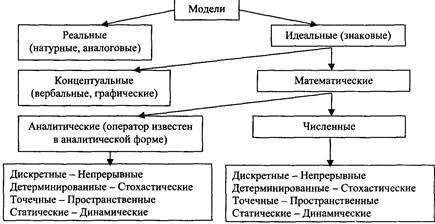
Рис. 10. Схематическая классификация моделей
По типу реализации различаются реальные и знаковые модели. Реальная модель отражает существенные черты оригинала уже по самой природе своей физической реализации (аквариум как модель природных водоемов).
Знаковая модель представляет собой условное описание системы-оригинала с помощью данного алфавита символов и операций над символами, в результате чего получаются слова и предложения некоторого языка, которые с помощью определенного кода интерпретируются как образы некоторых свойств элементов системы-оригинала и связей между ними.
Концептуальная модель представляет собой несколько более формализованный и систематизированный вариант традиционного естественнонаучного описания изучаемой экосистемы, состоящей из научного текста, сопровождаемого блок-схемой системы, таблицами, графиками и прочим иллюстративным материалом.
При количественном изучении динамики экосистем гораздо более эффективны методы математического моделирования. Если найдено точное аналитическое выражение, позволяющее для любых входных функций и начальных условий непосредственно определять значение переменных состояния в любой нужный момент времени, то модель принято называть аналитической. В то же время, если совокупность уравнений и неравенств непротиворечива и полна, то нередко удается найти алгоритм численного решения этих уравнений на ЭВМ. Такие модели называются численными, или имитационными.
В зависимости от степени определенности предсказания модели делятся на детерминированные и стохастические (вероятностные). В детерминированной модели зна-
чения переменных состояния определяются однозначно (с точностью до ошибок вычисления). Стохастическая модель для каждой переменной дает распределение возможных значений, характеризуемое такими вероятностными показателями. По характеру временного описания динамики переменных состояния различаются дискретные и непрерывные модели. Дискретная модель описывает поведение системы на фиксированной последовательности моментов времени t 0 ti ... /, ... tn , тогда как в непрерывной модели значения переменных состояния могут быть рассчитаны для любой точки t рассматриваемого интервала.
Модели, в которых пространственное строение экосистемы не рассматривается, принято называть моделями с сосредоточенными значениями (или точечными моделями), в отличие от моделей с распределенными значениями, в которых переменные состояния Xi зависят не только от времени, но и от пространственных координат (одной или нескольких).
4. Общая схема системного подхода к изучению экосистем
Изучение экосистемы оказывается наиболее эффективным, когда все методы -наблюдение, эксперимент, моделирование, интегрируются в едином процессе экологического исследования на основе системного подхода.
Этапы изучения экосистем схематически показаны на рисунке 11.
5. Факторы, действующие в экосистемах. Помехи в экосистемах Экологический фактор - одно из наиболее общих и чрезвычайно широких понятий экологии. Экологические факторы делятся на внешние (экзогенные) и внутренние {эндогенные) по отношению к данной экосистеме.
Второй распространенный классификационный принцип - это деление факторов на биотические и абиотические.
Широкое использование в экологической литературе находит классификация факторов в основном по отличительным свойствам экосистемы и внешней среды, специфическое влияние которых они характеризуют (метеорологические (климатические), геологические, гидрологические, миграционные (биогеографические), антропогенные, эдафические, и биотические.
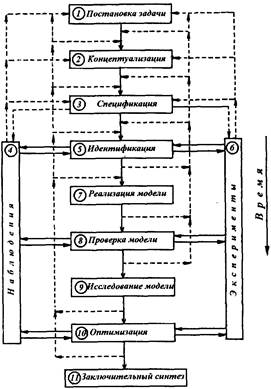
Рис. 11. Этапы изучения экосистем
Важным классификационным показателем служит характер временной динамики экологических факторов, в особенности наличие или отсутствие ее периодичности (суточной, лунной, сезонной, многолетней).
Особая роль среди экологических факторов принадлежит суммативным (аддитивным) факторам, характеризующим численности, биомассы или плотности популяций организмов, а также запасы или концентрации различных форм вещества и энергии, временные изменения которых подчиняются законам сохранения. Подобные факторы называются ресурсами (ресурсы тепла, влаги, органической и минеральной пищи). В отличие от них такие факторы, как интенсивность и спектральный состав радиации, уровень шума, окислительно-восстановительный потенциал, скорость ветра или течения, размер и форма пищи и т. д., которые также сильно влияют на организмы, относятся к категории условий.
По степени воздействия на организмы экологические факторы далеко не равносильны, Вследствие этого в экосистемах разного типа некоторые факторы выделяются как наиболее существенные, или императивные.
6. Пространство экологических факторов
Каждому экологическому фактору можно сопоставить математическую переменную, принимающую значение на некоторой шкале. Чтобы охарактеризовать множество всевозможных комплексов экологических факторов, получающихся при различных значениях каждого из них вводят понятие пространства экологических факторов - евклидово пространство, координаты которого сопоставлены ранжированным экологическим фактором. Для обозначения экологического пространства используют запись вида Е = {( xh х2 , ..., х„, ...)}, означающую, что множество Е состоит из всевозможных последовательностей вида ( xj , х2 .....х„, ...).
Таким образом, каждой конкретной комбинации значений экологических факторов X ] = Xi *. х2 = Х2*, ... , х„ = х„*, ... соответствует точка экологического пространства Е координатами ( xi *, x 2 *,..., х„* ...).
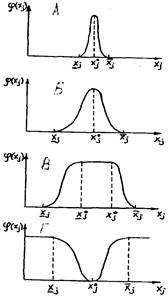
Рис. 12. Биологическая кривая
Хотя число экологических факторов потенциально неограниченно и, следовательно, бесконечны размерность экологического пространства Е, в действительности удается вылепить конечное число факторов (п), с помощью которых можно объяснить состояние организма в данный момент. Остальные, не вошедшие в число указанных факторов, не оказывают определяющего воздействия на изучаемый показатель. Их влияние можно рассматривать как некоторый «экологический шум-», накладывающийся на действие императивных факторов. Это позволяет от пространства Е с бесконечным числом измерений перейти к п-мерному подпространству Е„. Математически это можно записать следующим образом:
f (Xi, X2 ,..., X л , X n _i,...) = f (Хь Хг,..., Х„) + „+1, где f (x) - функция отклика некоторого показателя жизнедеятельности особей рассматриваемой популяции на экологические факторы; Хь Х2 , ..., Х„ -экологические факторы, Е„+ | - «экологический шум», вносящий незначительное возмущение в действие императивных факторов.
В типичных случаях график частной функции отклика на изменение фактора xs (i =1, ...п) имеет форму выпуклой кривой, монотонно возрастающей от минимального значения фактора х, (нижний предел толерантности) до максимума (или плато мак-
симальных значений) при оптимальных значениях фактора![]() , и монотонно убывающей, с приближением к максимальному значению (верхний предел толерантности)
(рисунок 12). Интервал
, и монотонно убывающей, с приближением к максимальному значению (верхний предел толерантности)
(рисунок 12). Интервал![]() , ограниченный минимальным и максимальным значениями i-
, ограниченный минимальным и максимальным значениями i-
того фактора, называется интервалом толерантности
по данному фактору, а точка (или интервал)![]() , в которой достигается максимальное значение показателя, называется точкой (интервалом или зоной) оптимума
по данному фактору. Виды, имеющие узкий интервал толерантности, называются стеноэками,
а виды с широким интервалом толерантности - эвриэками
по данному фактору.
, в которой достигается максимальное значение показателя, называется точкой (интервалом или зоной) оптимума
по данному фактору. Виды, имеющие узкий интервал толерантности, называются стеноэками,
а виды с широким интервалом толерантности - эвриэками
по данному фактору.
7. Основные представления о скоростях роста популяции
Скорость характеризует быстроту изменения чего-либо во времени. Так, число особей, родившихся за год, — это скорость рождаемости. Если N представляет собой число организмов, a t — время, то ДМ - изменение числа организмов, AN /At = средняя скорость изменения числа организмов за определенное время.
Часто необходимо знать не только среднюю скорость, но и теоретическую мгновенную скорость в тот или иной момент времени; иными словами, такую скорость, когда At стремится к нулю. В математическом анализе в тех случаях, когда рассматривают мгновенную скорость, Л заменяют буквой d . В этом случае предыдущее выражение приобретает вид: dNIdt = скорость изменения числа организмов за единицу времени в некоторый момент, dNI ( Ndt ) = скорость изменения числа организмов в расчете на 1 особь в единицу времени в некоторый момент.
Если среда не налагает никаких ограничений (пространство, пища и другие организмы не оказывают лимитирующего действия), удельная скорость роста (скорость роста популяции в расчете на 1 особь) для данных условий становится постоянной и максимальной. Значение скорости роста в этих благоприятных для популяции условиях представляет собой единственный показатель наследственно обусловленной способности популяции к росту. Этот показатель обозначают символом г.
dN / dt = rN ; r = dNI ( Ndt ).
Параметр г можно рассматривать как коэффициент мгновенного роста популяции. Когда устанавливается стационарное и стабильное возрастное распределение, удельную скорость роста называют внутренней скоростью естественного роста или Гтт- Максимальное значение г часто называют биотическим или репродуктивным потенциалом. Разницу между биотическим потенциалом и скоростью роста, наблюдаемой в реальных полевых или лабораторных условиях, часто используют как меру сопротивления среды, которая характеризует сумму всех лимитирующих факторов среды, препятствующих реализации биотического потенциала.
8. Кривые роста популяции. Характер увеличения численности популяции может быть различным, и в связи с этим выделяют различные типы роста популяции. По форме кривых роста можно выделить два основных типа: рост, описываемый J-образной и S -образной, или сигмоидной, кривой. В соответствии с особенностями разных организмов и условий среды эти кривые могут усложняться, модифицироваться или сочетаться различным образом. При J-образной кривой плотность быстро возрастает по экспоненте, но затем, когда начинают действовать сопротивление среды или другой лимитирующий фактор (обычно более или менее резко),рост быстро прекращается. Этот тип роста описывается экспоненциальным уравнением: dN / dt = rN .
При описываемом сигмоидной кривой типе роста популяция вначале увеличивается медленно (фаза положительного ускорения или становления), затем быстрее (возможно, приближаясь к логарифмической фазе), но вскоре под влиянием сопротивления среды рост постепенно замедляется (фаза отрицательного ускорения); в конце концов будет достигнуто равновесие, которое и сохраняется. Такой w, роста можно описать
логистическим уравнением:
Верхний предел увеличения![]() численности популяции, соответствующий константе К,
называется верхней асимптотой
сигмоидной кривой. Ее называют также преде-
численности популяции, соответствующий константе К,
называется верхней асимптотой
сигмоидной кривой. Ее называют также преде-
лом роста. При J-образном типе роста равновесие не достигается.
МОДЕЛЬ ЭКСПОНЕНЦИАЛЬНОГО РОСТА Мальтуса - одна из первых моделей динамики роста популяций, предложенная Т. Мальтусом (Т. Maltus) в 1799 г., в рамках которой динамика численности или плотности популяции N(t) описывается уравнением:
dN / dt = ( B - D )- N , или N ( t ) = N 0 exp(r t ), где В - коэффициент рождаемости; D - коэффициент смертности популяции, г = В - D (скорость роста популяции; подчеркнем, что для экспоненциального роста г = const ).
МОДЕЛЬ ОГРАНИЧЕННОГО РОСТА ПОПУЛЯЦИИ в 1825 г. предложил Б. Гомпертц (B.Gomperz), введя в уравнение Мальтуса следующую зависимость для разницы между коэффициентами рождаемости и смертности:
r ( N ) = rln ( N / K )/ lnK , где К- предельное значение характеристики популяции, которое может быть достигнуто при ее росте (г = const 0).
МОДЕЛЬ ЛОГИСТИЧЕСКОГО РОСТА Ферхюльста - Пирла Эмпирические исследования роста целого ряда популяций показали что «насыщение» (достижение порогового значения К) происходит гораздо раньше, чем это следует из модели Гомперт-ца. Модель Ферхюльста-Пирла - уравнение динамики численности или плотности популяции при условии ограниченного роста
r ( N ) = r -( K - N )/К или r ( N ) = ( a - b - N ). где а = r , b = г/К 0 - параметры линейной зависимости г от N.
Уравнение Ферхюльста-Пирла может быть записано и в более общем виде (не только линейная зависимость г от N ).
HN ) = r -[ l -(9 ( N ) L где cp ( N ) - зависимость интенсивности лимитирования от численности или плотности популяции. Логистическое уравнение с такой функцией (ср (N) иногда называют ОБОБЩЕННЫМ УРАВНЕНИЕМ РОСТА Ричардса. 9. Моделирование динамики сообществ
МОДЕЛИ С ЗАПАЗДЫВАНИЕМ - класс аналитических моделей, учитывающих наблюдающееся практически в каждой популяции запаздывание реакции организмов (рост смертности или замедление размножения) на изменения факторов окружающей среды. С учетом запаздывания в воспроизводстве потомства уравнение логистического роста примет следующий вид:
dN ( t ) / dt = r - N ( t )- [ K - N ( t - t ) J /К, где dN ( t )/ dt - скорость изменения численности в момент времени t N ( t ) численность популяции в момент времени t , N ( t - т) - численность популяции в момент времени ( t - т). МОДЕЛЬ КОНКУРЕНЦИИ Лотки-Вольтерра dNj / dt = n - Nr [( K ,- N ,- a ,2- N 2 )/ Ki ] dN 2 / dt = r 2 - N 2 - [( K 2 - N 2 - a 21 • N ,)/ K 2 ] и МОДЕЛЬ СИСТЕМЫ «ХИЩНИК-ЖЕРТВА» dN / dt = N ,- ( r , - b - N 2 ) dN 2 / dt = N 2 -( hb - N ,- m ), где N ,{ t ) - плотность популяций i в момент времени t (в системе «хищник-жертва» i = 1 - «жертва»; i = 2 - «хищник») г, - скорость экспоненциального роста популяций; А, -максимально допустимая плотность популяции (емкость экологической ниши); а$ - коэффициенты конкуренции; m - коэффициент естественной смертности хищников Ь - коэффициент хищничества к 1 - доля энергии содержащейся в биомассе жертвы которую хищник расходует на воспроизводство
МОДЕЛЬ ТОМА И ДЖЕРРИ Маргалефа - представления о параллельном коэво-люционном развитии системы с сопоставимым уровнем организации хищника и жертвы: «как хищник, так и жертвы оказываются включенными в эволюционный процесс, ведущий к увеличению размеров всех его участников. Во всех описанных до сих пор случаях взаимодействующие популяции животных и растений длительное время эволюционировали совместно и, таким образом, выработали стратегии не только для выживания, но и для взаимной выгоды.
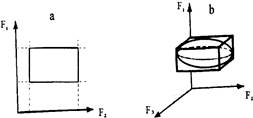
Рис. 13. Модели фундаментальной ниши Хатчинсона: а - двумерная экологическая ниша; b - трехмерная экологическая ниша
10. Модель фундаментальной ниши
МОДЕЛЬ МНОГОМЕРНОЙ (ФУНДАМЕНТАЛЬНОЙ) НИШИ Хатчинсона - представление фундаментальной ниши как некоторого гиперобъема в и-мерном пространстве абиотических и биотических факторов. Если на независимых осях-факторах отложить границы толерантности рассматриваемого организма (популяции) по отношению к этим факторам и восстановить из этих точек перпендикуляры то ограниченное ими пространство и будет определять гиперобъем экологической ниши данного организма (популяции). На рисунке 13 даны схемы представления двумерной и трехмерной экологических ниш.
Гомогенность пространства экологической ниши неодинакова, т.к. распределение вида вдоль градиентов среды колоколовидно. Иными словами, «...в пределах гиперобъема ниши существуют зоны, различающиеся по вероятности выживания в них». К тому же оси не могут быть независимы из-за совокупного действия природных факторов. Учет этого приводит к представлениям о «сферической» форме экологической ниши. В первом случае мы можем говорить о фундаментальной экологической нише, во втором - о реализованной.
11. Модель островной биогеографии Мак-Артура и Уилсона
В простейшей формулировке теория островной биогеографии состоит в том, что число видов на острове определяется равновесием между иммиграцией новых видов и вымиранием уже существующих. На рисунке 14 показаны четыре точки равновесия, соответствующие 1) небольшому удаленному острову, на котором предсказывается существование немногих видов; 2) большому, близкому к материку острову, который должен быть населен большим числом видов; 3) большой удаленный остров и 4) небольшой
близкий остров занимают промежуточное положение в отношении видового богатства.
Теория островной биогеографии дает основу для планирования резерватов, предназначенных для сохранения природного разнообразия.

Рис. 14. Теория островной биогеографии. Число видов на острове определяется равновесием между скоростями иммиграции и вымирания. Показаны четыре точки равновесия.
? Вопросы к семинару ?
1) Модели и моделирование в экологии, анатомия математических моделей.
2) Классификация моделей.
3) Общая схема системного подхода к изучению экосистем.
4) Факторы, действующие в экосистемах. Помехи в экосистемах.
5) Пространство экологических факторов.
6) Основные представления о скоростях роста популяции. Внутренняя скорость естественного роста.
7) Кривые роста популяции.
8) Моделирование динамики сообществ.
9) Модель многомерной экологической ниши Хатчинсона.
10) Модель островной биогеографии и ее прикладное значение.
Тема 5. Кибернетическая природа и стабильность экосистем
1. Кибернетическая природа экосистем
Помимо потоков энергии и круговоротов веществ, экосистемы характеризуются развитыми информационными сетями, включающими потоки физических и химических сигналов, связывающих все части системы и управляющих (или регулирующих) ею как одним целым. Поэтому можно считать, что экосистемы имеют кибернетическую (от греч. kybernetike — искусство управления) природу, хотя в отличие от созданных человеком кибернетических устройств ее управляющие функции сосредоточены внутри нее. Механические устройства, осуществляющие обратную связь, инженеры часто называют сервомеханизмами, тогда как биологи для живых систем используют термин гомеостатические механизмы. Кибернетическую природу экосистемы труднее выявить потому, что компоненты на экосистемном уровне связаны в информационные сети различными физическими и химическими агентами-«посредниками». В масштабе экосистемы эти слабые, но очень многочисленные связи энергии и химической информации были названы «невидимыми проводами природы».
Помимо системы обратной связи стабильность обеспечивается избыточностью функциональных компонентов. Например, если в сообществе имеется несколько видов автотрофов, каждый из которых характеризуется своим температурным диапазоном функционирования, то скорость фотосинтеза сообщества в целом может оставаться неизменной, несмотря на колебания температуры.
Степень стабильности, достигаемая конкретной экосистемой, зависит от ее истории, эффективности ее внутренних управляющих механизмов, характера среды на входе и от сложности экосистемы. Как правило, экосистемы имеют тенденцию становиться сложнее в благоприятной физической среде, чем в среде со стохастическими (непредсказуемыми) нарушениями на входе, например штормами.
Резистентная устойчивость — это способность экосистемы сопротивляться нарушениям, поддерживая неизменной свою структуру и функцию. Упругая устойчивость — это способность системы восстанавливаться после того, как ее структура и функция были нарушены. Оба типа стабильности взаимно исключают друг друга. Поскольку системе трудно одновременно развить оба типа устойчивости, обычно при благоприятных физических условиях среды экосистемы в большей степени проявляют резистентную, а в изменчивых условиях - упругую устойчивость. 2. Биологическая регуляция геохимической среды
ГИПОТЕЗА ГЕОМЕРИДЫ Беклемишева (1931) - одна из первых гипотез теоретической глобальной экологии. Геомерида - весь живой покров Земли, рассматриваемый как целостная иерархическая система, миллионы лет пребывающая в состоянии динамического устойчивого равновесия. Если биосфера - высший биотоп, то Геомерида - высший биоценоз.
ГИПОТЕЗА ГЕИ Лавлока-Маргулис (1979) - представление о биологическом контроле на биосферном уровне факторов абиотической среды и существовании
сложной, живой, саморегулирующейся системы поддержания на Земле условий благоприятных для жизни.
Известно, что абиотическая среда контролирует деятельность организмов. Организмы в свою очередь влияют на абиотическую среду и контролируют ее самыми разнообразными способами. Организмы постоянно изменяют физическую и химическую природу инертных веществ, отдавая в среду новые соединения и источники энергии. Это распространение биологического контроля на глобальный уровень стало основой гипотезы Геи (Гея — древнегреческая богиня Земли). Лавлок и Маргулис рассматривают сложную сеть микроорганизмов «коричневого пояса» как тонкую регулирующую систему, функционирующую по принципу хемостата, которая поддерживает пригодные для жизни условия. Эта регулирующая система («Гея») делает Землю сложной, но единой кибернетической системой.
ГИПОТЕЗА БИОТИЧЕСКОЙ РЕГУЛЯЦИИ Горшкова - представления о биотическом механизме регуляции окружающей среды на основе высокой степени замкнутости круговорота углерода. Потоки синтеза и разложения органических веществ скор-релированы с точностью 10~7 . Отношение потока отложения органического углерода к потоку его синтеза характеризует разомкнутость круговорота веществ. Естественная разомкнутость имеет значение порядка 104 , которое поддерживается с относительной точностью порядка 1(Г3 . Скоррелированность потоков синтеза и распада с указанной точностью доказывает наличие биологической регуляции окружающей среды, ибо случайная связь величин с такой точностью в течение миллионов лет невероятна.
3. Стабильность экосистем
Поскольку стабильные экосистемы (дождевой лес, коралловые рифы) обладают высоким видовым разнообразием, возникло искушение сделать вывод, что разнообразие повышает стабильность. Однако многочисленные виды, вступая друг с другом в конкурентную борьбу, могут дестабилизировать экосистему. Поэтому в природе нигде и никогда не достигается максимальное теоретическое разнообразие. В тех случаях, когда разнообразие высоко, в среднем, по-видимому, достигается 80 % максимального биоразнообразия.
Для того чтобы разнообразие внутри одного местообитания или типа сообщества не путать с разнообразием ландшафта или региона, который содержит смесь местообитаний, Уиттэкер (Whittaker) предложил следующие термины: 1) а-разнообразие для разнообразия внутри местообитания или внутри сообщества; 2) Р-разнообразие для разнообразия между местообитаниями и 3) у-разнообразие для разнообразия в обширных регионах биома, континента, острова и т. д.
К элементам структурного разнообразия в экосистемах относят:
1. Стратификационные принципы (вертикальная слоистость, ярусность растительного покрова и структура почвенных профилей); 2. Зональность (горизонтальная разобщенность, вертикальная поясность в горах или в литоральной зоне); 3. Характер активности (периодичность); 4. Структура пищевой сети (сетевая организация пищевых цепей); 5. Репродуктивные системы (ассоциации родителей и потомства, клоны растений и т. д.); 6. Социальные структуры (стада и табуны); 7. Системы взаимодействия (возникают в результате конкуренции, антибиоза, мутуализма и т. д.); 8. Стохастические структуры (возникают в результате действия случайных сил).
Увеличение разнообразия обусловлено также «краевыми эффектами» — контактами между пятнами контрастирующих типов растительности или физическими местообитаниями.
Стабильность более тесно связана с функциональным разнообразием: гусеница и бабочка или лягушка и головастик играют значительно более разнообразные роли в сообществе, чем два вида гусениц или взрослых лягушек.
Генотипическое разнообразие остается скрытым, если характеризовать сообщество только на уровне видов. Классическая теория основывалась на том, что особи в популяции должны быть гомозиготными по аллелям, дающим наибольшую приспособленность:
![]() . Альтернативная теория «сбалансированного полиморфизма»
утверждает, что
. Альтернативная теория «сбалансированного полиморфизма»
утверждает, что
особи гетерозиготны по большинству локусов:. Справедливость этой теории
подтверждается современными биохимическими![]() методами, которые могут выявить скрытую генетическую изменчивость. В отсутствие такой генотипической изменчивости виды оказались бы неспособными адаптироваться к новым ситуациям и, следовательно, должны были бы вымереть в изменяющейся среде.
методами, которые могут выявить скрытую генетическую изменчивость. В отсутствие такой генотипической изменчивости виды оказались бы неспособными адаптироваться к новым ситуациям и, следовательно, должны были бы вымереть в изменяющейся среде.
Биологическое разнообразие животных, растений и микроорганизмов представляет собой фактор фундаментальной важности для выживания человечества. На протяжении истории уменьшение видового и генетического разнообразия приносило кратковременную выгоду человеку при ведении лесного и сельского хозяйства.
Сейчас утверждается, что биота управляет окружающей средой, обеспечивая оптимальные условия для своего существования. Чтобы не допустить глобальной экологической катастрофы, необходимо обеспечить разнообразие в биосфере. Подсчитано, что при исчезновении 1/5 части видов растений и животных в биосфере начнутся цепные реакции, которые невозможно будет остановить. В этой связи необходимы финансовые затраты на сохранение видов растений и животных. Даже определена щена вида», которая рассчитана по следующей формуле:
В денежном выражении![]() (по данным 1994 г.) это составило 13 трлн. $ / 0,4-0,5 млн. видов = 32,5 млн. $; причем числитель этой дроби постоянно увеличивается, а знаменатель уменьшается, т.е. стоимость вида растет чрезвычайно быстро.
(по данным 1994 г.) это составило 13 трлн. $ / 0,4-0,5 млн. видов = 32,5 млн. $; причем числитель этой дроби постоянно увеличивается, а знаменатель уменьшается, т.е. стоимость вида растет чрезвычайно быстро.
? Вопросы к семинару ?
1) Кибернетическая природа экосистем и социальных систем. Типы устойчивости экосистем.
2) Биологическая регуляция геохимической среды, гипотеза Геомериды, гипотеза Геи, гипотеза биотической регуляции Горшкова.
3) Стабильность экосистем: структурная, функциональная, генетическая.
4) Озабоченность исчезновением видов и утратой биоразнообразия.
Тема 6. Надежность биосферы и техносфера
1. Надежность экосистем, биоценозов и биосферы
Любую экосистему можно определить как систему самовоспроизводящихся популяций, занимающих единый ареал обитания и выполняющих две основные функции. Первая функция - давление жизни - обеспечение прироста численности популяций или
общей биомассы экосистемы V (продуктивность данной экосистемы):. Вто-
рая функция - кондиционирование или очистка среды обитания от побочных![]() продуктов W:
продуктов W:![]() -0
(i = 1, 2, ..., N). Очевидно, чем больше скорость утилизации отходов, тем лучше условия для прироста биомассы, т.е. тем сильнее будет давление жизни.
-0
(i = 1, 2, ..., N). Очевидно, чем больше скорость утилизации отходов, тем лучше условия для прироста биомассы, т.е. тем сильнее будет давление жизни.
Оптимальным, по крайней мере для замкнутых экосистем о постоянный местообитанием, будет неравенство dV/dt = - dW/dt, которое соответствует поддержанию экосистемы в состоянии «максимальной чистоты», когда кондиционирующая мощность экосистемы превосходит ее продуктивность.
Биогеоценоз как самоорганизующаяся система, состоящая из нескольких (
N
)
экосистем, может достигнуть стабильности и замкнутости при условии![]()
когда скорость прироста биомассы в каждой экосистеме и популяции равна скорости образования побочных продуктов в режиме их утилизации.
При этом должно соблюдаться соотношение![]() , где
, где
п - численность каждой из N популяций (i = 1,2,..., N слагающих биосферу; Gj и D| - скорости размножения и гибели особей, входящих в эти популяции; U , - параметр конкуренции особей I -ой популяции между собой; уу - представленный в виде матрицы параметр, характеризующий взаимную утилизацию отходов организмами i -ой и j -ой популяций.
Данное соотношение - условие стабильного функционирования совокупности экосистем, состоящей из большого числа видов, в том числе и всей биосферы.
? Вопросы к семинару ?
1) Давление жизни и жизнеспособность популяций.
2) Надежность экосистем, биоценозов и биосферы.
3) Факторы, лимитирующие техногенез. Ограничения численности популяций и направления эволюции. Роль технологий в приросте человечества.
4) Этапы эволюции человечества.
5) Ноогенная техносфера. Стратегия выживания человечества.
Тема 7. Объяснение и прогнозирование в экологии
При исследовании простых систем (например, в классической физике) функции объяснения и предсказания совмещаются в рамках одного закона (Закон всемирного тяготения Ньютона и т.п.) Для сложных свойств сложных систем одна модель (один закон) будет не в состоянии одновременно удовлетворительно выполнять как объяснительную, так и предсказательную функции.
1. Логическая структура научного объяснения и предсказания
Процесс объяснения заключается в том, что некоторые явления или свойства сложных систем пытаются подвести под заранее установленные и принятые в данной теории законы и гипотезы (дедуктивное объяснение). Методы индуктивного объяснения, связанны с выдвижением статистических гипотез и получением статистических описаний для объясняемого явления. В этот класс следует отнести методы экстраполяции, адаптивных оценок и аналогий.
Методы предсказания также делятся на дедуктивные (в количественном прогнозировании это - имитационные модели) и индуктивные (классический регрессионный анализ). Различия этих процессов объяснения и предсказания заключаются в том, что предсказание имеет положительную направленность во времени (относится к настоящему или будущему), а объяснение - отрицательную (к настоящему или прошлому).
Научное прогнозирование - это специальное исследование, имеющее свою методологию и технику, проводимое в рамках управления, с целью повышения уровня его обоснованности и эффективности.
Исследование будущего разделяется на два качественно различных направления:
- Поисковое прогнозирование - это анализ перспектив развития существующих тенденций на определенный период и определение на этой основе вероятных состояний объектов управления в будущем при условии сохранения существующих тенденций в неизменном состоянии или проведения тех или иных мероприятий с помощью управленческих воздействий;
- Нормативное прогнозирование представляет собой попытку рационально организованного анализа возможных путей достижения целей оптимизации управления.
2. Имитационные модели глобальных процессов
ИМИТАЦИОННЫЕ МОДЕЛИ ГЛОБАЛЬНЫХ ПРОЦЕССОВ в биосфере - модели для описания изменений компонент экосистем (биогеохимически. циклов) под воздействием антропогенных факторов в масштабе биосферы.
Одной из первых глобальных моделей изменения биосферы атмосферы и климата была модель В.А. Костицына (1935). Моделями биогеохимических циклов занимался Крапивин. Создание ЭВМ позволило рассмотреть очень сложные проблемы, важные для всего человечества. Возникала наука «глобалистика», основанная на изучении с помощью моделирования на ЭВМ общепланетарных проблем, глобальных проблем человечества. В конце 70-х - начале 80-х годов в ВЦ АН СССР под руководством Н.Н. Моисеева была создана версия глобальной модели биосферы названная «Системой Геи». С ее помощью был проанализирован сценарий «локального ядерного конфликта», описан эффект «ядерной зимы» и дан прогноз глобальных изменений в биосфере. При моделировании функционирования биосферы выяснилось, что после крупномасштабных воздействий биосфера никогда не возвращается в исходное состояние. Каждый раз это будет новая биосфера, и ее параметры, как правило, исключают возможности дальнейшего развития человека.
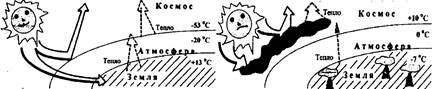
Рис. 15. Иллюстрация эффекта ядерной зимы (температура указана для поверхности почвы, средних и верхних слоев атмосферы)
3. Модели Римского клуба
Методологической основой построения прогнозов в глобальном масштабе стали методы математического моделирования и, прежде всего, методы системной динамики Дж. Форрестера. Он построил примитивную, но достаточно всеобъемлющую математическую модель, которая могла бы грубо имитировать развитие мировой ситуации с помощью пяти основных взаимозависимых переменных: численности населения, объема капиталовложений, использования невозобновимых ресурсов, производства продовольствия и загрязнения среды.
Модель «Мир-1» состояла из 42 нелинейных уравнений, описывающих взаимосвязь между выбранными переменными. Первая модель: увеличивается население - ускоренный рост всех остальных показателей. Прогноз: экологическая катастрофа между 30 и 50 годами 21 века.
Вскоре профессор Д. Медоуз с группой ученых усовершенствовал модель мира, получив модель «Мир-2», или «модель стабилизации общества»: после 1975 г. прирост населения должен быть равен нулю, капиталовложения в промышленность - равняться амортизации, вводится повторное использование ресурсов и внедрение безотходных технологий. Прогноз: стабилизация, при которой уровень производства на душу населения будет в три раза превышать среднемировой уровень 1970 года.
Третья модель «Мир-3»: те же меры, что и в модели стабилизации, но не с 1975, а с 2000 года. Прогноз: равновесие не достигается. Недостаток продовольствия будет ощущаться ранее 2100 года.
К середине 1980-х годов имелось более 15 глобальных прогнозов, получивших название моделей мира. Самые известные и наиболее интересные из них - это Мировая динамика Дж. Форрестера, Пределы роста Д. Медоуза с соавторами, Человечество у поворотного пункта М. Месаровича и Э. Пестеля, Латиноамериканская модель Баричо-ле А. О. Эрреры (ЛАММ), Будущее мировой экономики В. Леонтьева, Мир в 2000 году. Доклад президенту Британская SARUM-модель, японская модель FUGI и мировая модель ООН (UNWM) и другие.
4. Принципы устойчивого развития
Основные проблемы, стоящие на пути достижения цели устойчивого развития, таковы:
1. Рост населения; 2. Проблема производства продуктов питания, защита ресурсов и окружающей среды; 3. Сохранение почвы; 4. Охрана водных ресурсов Земли; 5. Защита лесов; 6. Защита атмосферы Земли; 7. Управление отходами, образуемыми в
процессе человеческой деятельности; 8. Эффективное использование энергии; 9. Развитие промышленности и экологизация технологий; 10. Устойчивость экосистем; 11. Сохранение биологического разнообразия; 12. Ответственность и значение отдельных личностей за экологически обоснованный выбор и достижение поставленных целей.
Концепция устойчивого развития интегрирует в себе экологическую экономическую и социальную сферы и является политическим документом для успешной реализации которого необходимо последовательное проведение демократизации управления и соблюдение следующих принципов:
- Принцип иерархической организации. Каждый уровень (крупные и средние города, области, автономии; регионы; государства, федерации; планета Земля) иерархической организации территории решает задачи в пределах своей компетенции, за счет собственных сил и средств, оказывая методическую и консультативную помощь нижележащим уровням и представляя интересы последних перед более высокими уровнями.
- Принцип единства целей. Деятельность всех уровней иерархии объединяется единством целей: обеспечение мира и безопасности; рациональное экологически сбалансированное природопользование; рациональное использование естественных и антропогенно-измененных ландшафтов, охрана разнообразия растительного и животного мира, эталонных природных систем, реконструкция нарушенных ландшафтов.
- Принцип последовательной экологизации всех сфер жизнедеятельности. Ресурсы каждой территории находятся в собственности и распоряжении ее населения и используются для удовлетворения основных материальных, духовных, эстетических потребностей, обеспечения здоровья населения, полноценного его воспроизводства и достижения целей устойчивого развития.
- Принцип «управление - для населения». Управление территориями строится на принципе передачи местным органам максимально возможных, а центральной власти -минимально необходимых полномочий в принятии решений, а также на основе достижения общественного согласия по наиболее существенным вопросам, затрагивающим интересы всего населения или отдельных групп.
- Принцип единого контроля и доступности информации. Население имеет право получать любую информацию, касающуюся экологической обстановки; органы управления не могут препятствовать получению и распространению информации и участию населения в решении проблем устойчивого развития территории. Любой род деятельности открыт для служб экологического контроля всех уровней.
- Принцип финансирования программ устойчивого развития. Этапность выполнения программ устойчивого развития территорий любого масштаба следует напрямую связать с выделением в защищенных разделах бюджетов территорий (страны, области, города) отдельной строкой расходов на охрану окружающей природной среды в объемах: I этап - не менее 3 % расходной части бюджета; II этап - не менее 5 % расходной части бюджета; III этап - не менее 7 % расходной части бюджета;
- Принцип осуществимости программ устойчивого развития. На каждом иерархическом уровне для достижения устойчивого развития необходима реализация следующих систем обеспечения такой Программы: законодательно-правовой и нормативной; экономической; контрольно-информационной (мониторинговой); научной; просветительно-образовательной; материально-технической и кадровой; организационной.
? Вопросы к семинару ?
1) Научное объяснение и предсказание.
2) Глобалистика и каковы ее задачи. Имитационные модели глобальных процессов в биосфере.
3) Модель «ядерной зимы».
4) Модели Римского клуба.
5) Модели мира разных авторов.
6) Основные проблемы, стоящие на пути достижения устойчивого развития.
7) Принципы устойчивого развития.
Библиографический список
Основная литература
Волкова В. П. Основы теории систем и системного анализа / В. П. Волкова, А. А. Денисов. СПб.: СПб. ГТУ, 1997.
Корогодин В. И. Информация и феномен жизни. - Пущино, 1991.
Маргалеф Р. Облик биосферы. М.: Наука, 1992.
Марчук Г. И. Математическое моделирование в проблеме окружающей среды. М: Наука, 1982.
Математические модели в экологии и генетике. М., 1981.
Михайловский Г. Е. Жизнь и ее организация в пелагиали Мирового океана. М.: Наука, 1992.
Мозговой Д. П. Информационные поля и поведение млекопитающих / Д. П. Мозговой, Г. С. Розенберг, Э. Д. Владимирова. - Самара: Изд-во Самар. ун-та, 1998.
Моисеев Н. Н. Математические задачи системного анализа. М.: Наука, 1981.
Моисеев Н. Н. Человек и ноосфера. М.: Молодая гвардия, 1990.
Одум Ю. Экология: В 2 т. Т. 1: Пер. с англ. М.: Мир, 1986.
Петросян Л. А. Введение в математическую экологию / Л. А. Петросян, В. В. Захаров. -Л.: Изд-во Ленингр. ун-та, 1986.
Реймерс Н. Ф. Экология (теории, законы, правила принципы и гипотезы). М.: Россия Молодая, 1994.
Реймерс Н.Ф. Природопользование: Словарь-справочник. М.: Мысль, 1990.
Розенберг Г. С. Экология. Элементы теоретических конструкций современной экологии: Учеб. пособие / Г. С. Розенберг, Д. П. Мозговой, Д. Б. Гелашвили. - Самара: Самарский научный центр РАН, 1999.
Рубин А. Б. Термодинамика биологических процессов. М.: Изд-во Моск. ун-та, 1984.
Свирежев Ю. М. Нелинейные волны, диссипативные структуры и катастрофы в экологии. - М.: Наука, 1987.
Смит Дж. Модели в экологии. - М.: Мир, 1987.
Федоров В. Д. Экология / В. Д. Федоров, Т. Г. Гильманов. М.: Изд-во МГУ, 1980.
Чистохвалов В. Н. Охрана окружающей среды: прогнозирование и методы принятия решений. М.: Изд-во РУДН, 1992.
Дополнительная литература
Аптер М. Кибернетика и развитие. М.: Мир, 1970. Блюменфельд Л. А. Проблемы биологической физики. М.: Наука, 1977. Бриллюэн Л. Наука и теория информации. М.: Гос.-изд. физ.-мат. лит., 1960. БудыкоМ. И. История атмосферы / М. И. Будыко, А. Б. Ронов, А. Л. Яншин. Л.: Гидрометеоиздат, 1985.
Вернадский В. И. Автотрофность человечества // Проблемы биогеохимии: Тр. биогеохим. лаборатории. М.: Наука, 1980. Вып. 16.
Вернадский В. И. Биосфера и ноосфера. М.: Наука, 1989.
Вернадский В. И. Биосфера. Статьи по биогеохимии // Избр. соч. М.: Изд-во АН СССР. 1960. Т. 5.
Винер Н. Кибернетика или управление и связь в животном и машине. М.: Сов. радио, 1968.
Горшков В. Г. Устойчивость биосферы и сохранение цивилизации / В. Г. Горшков, К. Я. Кондратьев, С. Г. Шерман // Природа. 1990. № 6.
Корогодин В. И. Теория информации и биология. - Препринт ОИЯИ, PI9-83-590, Дубна, 1983.
Миркин Б. М. Популярный экологический словарь / Б. М. Миркин, Л. Г. Наумова. М., 1999.
Миркин Б. М. Современная наука о растительности: Учеб. / Б. М. Миркин, Л. Г. Наумова, А. И. Соломещ. М.: Логос, 2001.
Пархоменко В. П. Ядерная зима / В. П. Пархоменко, А. М. Тарко // Экология и жизнь. 2000. № 3 .
Печчеи А. Человеческие качества. М.: Прогресс, 1980.
Поплавский Р. П. Термодинамика информационных процессов. М.: Наука, 1981.
Розенберг Г. С. Модели в фитоценологии. М.: Наука, 1984.
Свирежев Ю. М. Устойчивость биологических сообществ / Ю. М. Свирежев, Д. О. Логофет. М.: Наука, 1978.
Седов Е. А. Эволюция и информация. М.: Наука, 1976.
Серавин Л. Н. Теория информации с точки зрения биолога. Л.: Изд-во Ленингр. ун-та, 1973.
Тейяр де Шарден П. Феномен человека. М.: Прогресс, 1965.
Хотунцев Ю. Л. Человек, технологии, окружающая среда. М.: Устойчивый мир, 2001.
Чернавский Д. С. Синергетика и информация. М.: Знание, 1990.
Содержание
с.
Введение. Становление и развитие системных идей в экологии..................................3
Тема 1. Основы теории систем и системного анализа. Системы и закономерности их формирования и развития ..............................................................4
1. Система. Простые и сложные системы. Классификация систем.....................................4
2. Иерархия уровней организации..........................................................................................4
3. Иерархический, сетевой и реляционный подходы к анализу систем..............................6
4. Основные принципы системологии....................................................................................6
Тема 2. Элементы системного анализа в экологии и охране окружающей
природной среды .............................,........................................................................7
1. Структура экосистемы.........................................................................................................7
2. Гипотеза однонаправленности потока энергии.................................................................8
Тема 3. Экология биосферы (Вещественные, энергетические и информационные процессы в экосистемах) ........................................................................8
1. Типы земного вещества.......................................................................................................8
2. Основные экологические законы, связанные с веществом биосферы............................8
3. Главные биогеохимические циклы биосферы........................................................................9
4. Термодинамика экосистем: закон энтропии....................................................................11
5. Экологические законы, связанные с энергетическими потоками биосферы................11
6. Элементы биоэнергетики экосистем....................................................................................12
7. Концепция продуктивности...............................................................................................14
8. Концепция информации.....................................................................................................14
9. Информация в растительных сообществах......................................................................15
10. Информационные поля животных..................................................................................16
11. Информация и феномен жизни.......................................................................................16
Тема 4. Модели и моделирование в экологии .................................................................17
1. Модели.................................................................................................................................17
2. Анатомия математических моделей.................................................................................17
3. Классификация моделей....................................................................................................18
4. Общая схема системного подхода к изучению экосистем.............................................19
5. Факторы, действующие в экосистемах. Помехи в экосистемах....................................19
6. Пространство экологических факторов............................................................................20
7. Основные представления о скоростях роста популяции................................................21
8. Кривые роста популяции...................................................................................................21
9. Моделирование динамики сообществ..............................................................................22
10. Модель многомерной (фундаментальной) ниши Хатчинсона.....................................23
11. Модель островной биогеографии Мак-Артура и Уилсона...........................................23
Тема 5. Кибернетическая природа и стабильность экосистем ....................................24
1. Кибернетическая природа экосистем...............................................................................24
2. Биологическая регуляция геохимической среды.............................................................24
3. Стабильность экосистем....................................................................................................25
Тема 6. Надежность биосферы и техносфера ..................................................................26
1. Надежность экосистем, биоценозов и биосферы............................................................26
Тема 7. Объяснение и прогнозирование в экологии ......................................................27
1. Логическая структура научного объяснения и предсказания..............................................27
2. Имитационные модели глобальных процессов в биосфере.................................................27
3. Модели Римского клуба............................................................................................................28
4 Принципы устойчивого развития.......................................................................................28
Библиографический список...............................................................................................30