Морфология колонии и признаки морфофункциональной адаптации гидроида
СОДЕРЖАНИЕ: На основе изучения беломорских Obelia geniculata, культивируемых как в естественных, так и в лабораторных условиях, определены характерные морфологические признаки гидроидов этого вида, тесно связанные с адаптацией к жизни на сильном течении.obelia geniculata (l.) К существованию в местах с сильным течением
Н. Н. Марфенин, А. В. Хоменко
На основе изучения беломорских Obelia geniculata, культивируемых как в естественных, так и в лабораторных условиях, определены характерные морфологические признаки гидроидов этого вида, тесно связанные с адаптацией к жизни на сильном течении. Короткие слабо разветвленные побеги обладают небольшой парусностью. Короткие ножки гидрантов и слегка асимметричные гидротеки уменьшают биение гидрантов в струях воды, Ширококоничесние гидротеки прикрывают гидранты от прямого действия течения. Отсутствие перетяжек в основании побегов делает их менее гибкими. Уплощенные столоны способствуют лучшему креплению колоний к субстрату. Активная фрустуляция позволяет осваивать прилежащий субстрат даже на сильном течении.
The morphological characteristics of hydroids closely connected with the life adaptation in the strong stream were determined studying Obelia geniculata from the White Sea cultivated under either natural or laboratory conditions. Short and poorly branched shoots have low sail effect. The short stalks of the hydranths and slightly asymmetric hydrothecas decrease the hydranth vibration in the water streams. The wideconic hydrothecas protect the hydranths from direct stream action. The absence of annull of periderm makes stalks not so flexible. Better attachment to substrate is achieved by flatting of stolons. The active frusulation gives possibility to study the adjoined substrate even in the strong stream.
Настоящая статья продолжает серию работ по монографическому описанию отдельных видов гидроидов [3, 6—9], выполненных по единой схеме, позволяющей сравнивать эти виды по достаточно широкому ряду унифицированных показателей. Необходимость проведени собственных исследований продиктована фрагментарностью литературных сведений, а также малой их сопоставимостью из-за различий условиях наблюдений и опытов.
Наряду с изучением строения колоний у гидроидов нас интересовала морфофункциональная подоплека деления экологических ниш между разными видами. Поэтому мы провели подробное исследована морфологии колонии и некоторых аспектов биологии у нескольки видов, принадлежащих к роду Obelia. Это дало возможность глубже понять особенности и способы приспособления каждого вида к типичным для них условиям обитания и функциональное значение ряда морфологических признаков, некоторые из которых были важны только для систематики, а некоторые и вовсе ранее не рассматривались. В данной статье мы ограничиваемся описанием одного из этих видов — Obelia geniculata (L.).
Методика
Материал был собран на Белом море, а исследования проводили на Беломорской биостанции МГУ летом 1981 г. на живых колониях, которые выращивали н пластинах из оргстекла по методике, описанной ранее [I]. Пластины содержали ка в лаборатории, так и в море. Для описания строения колонии (расположения побегов, столонов, верхушек роста, гидрантов, гонангиев) применяли модифицированный метод картирования [4, 8].
Авторы благодарят своих коллег И. А. Косевича, Т. Н. Хоменко и М. Плетникову за участие в обсуждении полученных материалов
Результаты исследований
В естественных условиях колонии О. geniculata обычно встречаются на ламинариях и не производят впечатления густых зарослей. Не ветвящиеся короткие побеги кажутся расположенными редко на хорошо просматриваемой сквозь них гидроризе (рис. 1). Ветви гидроризь (столоны) расходятся лучами в разные стороны от центра колонии. От них, в свою очередь, не слишком часто отходят боковые ветви. Та кое строение колонии мы называем центробежным [8]. Перекрещиваясь друг с другом, столоны образуют негустую сеть на субстрате Из-за коротких междоузлии, укороченных ножек гидрантов, расположения всех гидрантов в одной плоскости побеги О. geniculata похож на побеги Dynamena pumila (L.). Сходство столь сильно, что различие часто становится очевидным лишь после изучения под микроскопом .Старейшие побеги, расположенные в центре колонии, отличаются от всех остальных своей большей длиной и разветвленностью. Площадь занимаемая средней по размерам колонией, равна примерно 100— 150 см2.
В лаборатории колонии выглядят несколько по-иному из-за более интенсивного ветвления побегов.
Молодые колонии появляются в условиях Белого моря на талломах ламинарии с конца мая — начала июня. Первые медузы О. geniculata были обнаружены в планктонных пробах лишь в конце июня;а последние — в начале сентября.
Нитевидная гидрориза представлена обычно одним или несколькими (до 5) основными столонами (т. е. столонами I порядка), отходящими от первоначального побега, а также многочисленными столонами II, III порядков и т. д. (см. рис. 1). Все они плотно прилегают к субстрату. Трубка столона на поперечном срезе слегка сплющена (рис. 2, ). По бокам вдоль всей трубки перисарк утолщен. Сверху это выглядит так, как будто вдоль трубки ценосарка с обеих сторон идут довольно широкие желтые полосы. В верхушке столона перисарк еще тонок, а ценосарк, занимая все пространство, выглядит, как широкое копье (рис. 2, Б); проксимальнее ценосарк сужается, занимая лишь часть полости по оси перисарка.
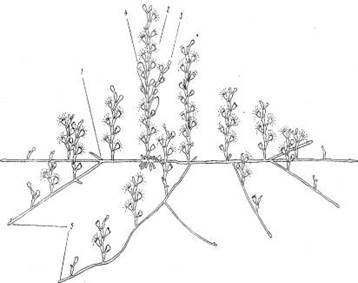
Рис. 1. Общий вид колонии ОЬеliа geniculata:
/ — гидрорйза, 2 — побег, 3 — боковая ветвь, 4 —гонангий, 5 — верхушка гидроризы
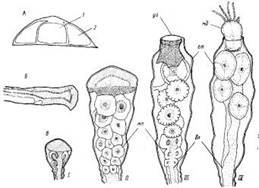
Рис. 2. Obelia geniculata. A—поперечный срез гидроризы; Б—верхушка гидроризы сверху; В — последовательные стадии развития гонангия; /—10 августа 1981 г.;//—через 50 ч после /; III—через 47 ч после //; IV—через 47 ч после ///: / — ценосаркальная полость, 2 — утолщение перисарка; бл — бластостиль, гт — гонотека, уг — устье гонотекн, мп — медузоидная почка, мд — медуза
*
Образование боковых ветвей гидроризы, т. е. столонов очередного порядка, происходит латерально на некотором удалении (4—12 мм и более) от верхушки столона обычно вблизи оснований побегов. Образование побегов происходит регулярно по мере роста столона, так что расстояние между побегами примерно одинаковое (3—3,5 мм). В выборке из 300 измерений все вариации расстояния между побегами составляли от 1 до 7 мм (рис. 3).
Растут столоны обычно прямолинейно либо по плавной дуге- Более резкие изменения в направлении роста столонов происходят лишь при встрече с препятствиями,
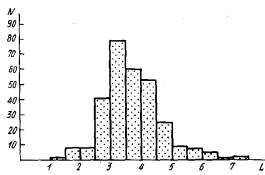
Рис. 3. Вариационный ряд величин расстояний между побегами на столонах. N—количество наблюдений; L—расстояние между побегами, мм
Побеги редко превышают в высоту 15—20 мм. Самый высокий из найденных нами достигал 28 мм. Ствол и боковые ветви имеют одинаковое строение и состоят из четких стереотипных междоузлии (рис. 4,4). Длина междоузлия (без ножки гидранта, в которую оно переходит) в среднем 0,7 мм (от 0,25 до 1,35 мм, выборка 471). Каждое междоузлие слегка расширяется в дистальном направлении, оставаясь уплощенным в плоскости побега, и дугообразно изгибается (радиус наибольшего изгиба междоузлия составляет 0,5 мм).
Междоузлия имеют проксимальную зону коль-чатости, которая состоит из 1—4 (обычно 2—3) четковидных вздутий. Кольчатая ножка гидранта как бы продолжает междоузлие. Она примерно в 3 раза тоньше (0,13 мм) наиболее широкой дистальной части междоузлия. Длина ножки обычно 0,25—0,45 мм. В ней насчитывается от 1 до 8 колец (вздутий).
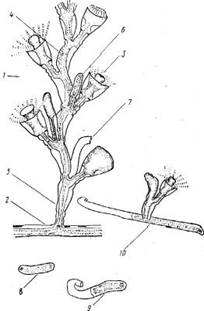
Рис. 4. Рост и развитие фрустул:
1 — побег, 2 — столон, 3 — гидрант, 4 — междоузлие, 5 — ножка побега, 6 — формирующая фрустула, 7— трубка перисарка после отделения фрустулы, 5 — осевшая фрустула, 9 — рост столона из фрустулы, 10 — образование побега
Очередное междоузлие, начинающееся рядом с ножкой гидранта, имеет изгиб в противоположном направлении, но при этом лежит в одной плоскости с предыдущим. Таким образом междоузлия поочередно отклоняются в обе стороны от оси побега на 20—30°. Самое первое (базальное) междоузлие побега почти перпендикулярно к столону и проксимальной зоны кольчатости не имеет. Иными словами, в месте соединения побега со столонами ясных перетяжек перисарка нет.
Первое междоузлие боковой ветви образуется в пазухе между гидрантом и стволом побега и изогнуто по направлению к стволу. Его ось ориентирована по отношению к оси побега под углом 50—60°. Плоскость боковой ветри не всегда совпадает с плоскостью ствола побега..
Кроме обычных боковых ветвей у О. geniculaia регулярно возникают многочисленные отростки, служащие для образования фрустул. Эти фрустулообразующие ветви закладываются так же, как и боковые ветви, — в пазухах между ножками гидрантов и стволом. Далее, однако, они не образуют характерных изогнутых междоузлии, а формируют слегка расширяющиеся и почти прямые трубки протяженностью 1—1,5 мм (см. рис. 4). Ценосаркальная верхушка фрустулооб-разующей ветви постепенно теряет связь с остальным ценосарком и продолжает расти вперед самостоятельно до тех пор, пока не коснется субстрата, и дальше растет уже на нем, формируя колонию, либо до тех пор, пока не отвалится вследствие хрупкости тонкого перисарка, остающегося проксимальнее фрустулы. В последнем случае фру-стула, падая на субстрат, прилипает к нему и в процессе дальнейшего роста образует колонию (рис. 4,70). Сначала в течение 4—6 суток растет столон, а затем вблизи его верхушки возникает зачаток побега.
Фрустула по форме и размерам напоминает плану лу, в отличие от которой не плавает, не сжимается и не изгибается, а, кроме того, покрыта перисарком. Длина фрустулы 0,5—0,6 мм, ширина 0,1 мм.
Фрустуляция у О. geniculata временами идет очень активно. Например, на побеге с 21 междоузлием было отмечено 10 фрустул, что составляло около 24 % длины всего ценосарка побега. В колонии часть ценосарка, идущая на образование фрустул, может достигать 19%.
Таким образом, гидроид размножается вегетативно в течение всего роста колонии, что играет важную роль в его экологии.
Гидранты типичной для Campanulariidae формы (рис. 4,3). В сжатом состоянии тело гидранта целиком помещается в гидротеке, прикрепляясь к ней шайбовидным расширением в базальной части на границе с ножкой. На дистальном конце тело гидранта слегка расширяется, давая один венчик из 26—30 шупалец, которые располагаются в два яруса, загибаясь по очереди то вверх, то вниз. Нижний ярус венчика, опираясь на край гидротеки, фиксирует положение гидранта. Все щупальца устроены одинаково и формируются одновременно. Их длина в расправленном состоянии 0,55—0,8 мм, диаметр 0,02—0,03 мм.
Колокольчатая (бокаловидная) гидротека слегка асимметрична — прилежащая к стволу сторона несколько короче, чем отстоящая, за счет чего гидротека в большей степени ориентирована вдоль ствола. Края гидротеки слегка отогнуты наружу. В колониях, росших в естественных условиях, высота гидротеки 0,48—0,76 мм (в среднем 0,65мм), а диаметр ее устья 0,46—0,70 мм (в среднем 0,64 мм). Показатель отношения высоты гидротеки к диаметру ее устья колеблется от 0,95 до 1,45 (в среднем 1,16). У колоний, которые культивировали в лаборатории, размеры гидротек более постоянны: высота 0,53—0,70 мм, ширина устья 0,53—0,70 мм при отношении высоты гидротеки к ее диаметру 0,98—1,01.
Продолжительность жизни гидранта обычно 7—10 суток (выборка равна 107), но бывает и дольше — до 16 суток. Затем гидрант за 8— 12 ч рассасывается. Пустая гидротека отваливается. Остается лишь ножка гидранта, на апикальном конце которой при благоприятных условиях вскоре (через сутки или более) начинается формирование следующего гидранта.
В гидротеке щупальца укладываются поверх булавовидного гипостома, заменяя отсутствующую крышечку гидротеки.
Питаются гидранты зоопланктом. Взрослые Copepoda, в том числе Harpacticoida, метанауплиусы усоногих, а также бентосные Amphipoda, нематоды, моллюски для гидрантов либо слишком велики, либо сильны и подвижны, либо обладают предохраняющими их шипами. Мы наблюдали, как гидранты заякоривали и заглатывали трохофор, велигеров, молодых хетогнат, планктонных гарпактицид Microsetella и небольших калянусов. Свежевылупившиеся науплиусы Artcmia salina представляют собой хороший лабораторный корм для О. geniculata. Гидранты заглатывают их обычно за 15—18 мин (от 7 до 32 мин при выборке 50). Время переваривания в четырех наблюдениях было соответственно 80,100, 130 и 195 мин.
Одновременно гидрант способен переваривать до трех науплиусов, которые могут быть проглочены один за другим. Продолжительность заглатывания науплиуса зависит от его ориентации по отношению к гипостому.
Сытый гидрант все равно заякоривает и убивает добычу, но спустя некоторое время отпускает ее со щупалец. Во время кормления у основания побегов может скапливаться большое количество убитых науп-лиусов. Таким образом, по пищевому поведению О. geniculata не отличается от других Campanulariidae [8].
У О. geniculata на Белом море формирование гонангиев продолжается с конца июня по конец августа, преимущественно на больших побегах колонии. В этот период они находятся в колонии на разных стадиях развития. Почка гонангия закладывается на прилежащей к стволу стороне ножки гидранта, ближе к ее основанию, и в процессе развития проходит ряд характерных стадий (рис. 2,5). У сформированного гонангия высота гонотеки достигает 3 мм, диаметр в наиболее широком месте 0,6 мм, диаметр устья 0,3 мм. Короткая ножка гонангия четковидна, диаметром около 0,3 мм. Различий между мужскими и женскими гонангиями не обнаружено.
Медузоидные почки (гонофоры) закладываются на бластостиле в проксимальной его части. Затем, по мере роста бластостиля, они продвигаются к устью гонотеки, одновременно дифференцируясь в маленьких медузок. Обычно бластостиль, продолжая расти, выталкивает верхних сформированных медузок через устье гонотеки. При этом рост происходит ритмичными пульсациями. Свернутая медузка втискивается в устье гонотеки, в котором бластостиль образует пробку. Пробка отжимается медузкой, образуя щель, через которую она выворачивается наружу (рис. 2, В, IV}. Медузки отделяются от бластостиля и, становясь свободными, уплывают. Иногда, впрочем, отделение медузки от бластостиля происходит еще внутри гонотеки. Тогда она переворачивается и присасывается к бластостилю ротовым стебельком; благодаря этому добирается до устья и выходит наружу.
Медузки с 27—34 щупальцами (у взрослых до 100 щупалец) уже могли проглотить свежевылупившегося науплиуса артемии. Более развитые медузы легко заякоривали и заглатывали планктонных гарпак-тицид Microsetella, молодых хетогнат, личинок и мелких калянусов.
Обсуждение результатов
А. А. Бялыницкий-Бируля [2] одним из первых обратил внимание на зависимость некоторых особенностей строения гидроидов от условий их обитания. Он нашел, что у литоральных колоний Campanularia integra гидротеки толстостенные, а у колоний из сублиторали — тонкостенные. Д.В. Наумов [10] в статье, посвященной влиянию движения воды на морфологию гидроидов, показал, что у литоральной формы С. integra, кроме того, более ясно выражена членистость ножек гидро-тек, что у Sertularella rugosa (L.) на литорали гидротеки также более толстые, прочные и менее морщинистые, а сами побеги менее разветвленные, чем у экземпляров с глубины 30 м. Д. В Наумов связал указанные различия в морфологии гидроидов с приспособлением их к жизни в местах, различающихся по подвижности воды. На литорали в прибойной зоне, по его мнению, важно как укрепление гидротек, так и увеличение гибкости побегов, которая, в частности, становится больше при наличии перетяжек на стволе и на ножках гидрантов.
Впечатляющие исследования Ридла [12—14] и Ведлера [15], выполненные под водой с применением легководолазного снаряжения, продемонстрировали зависимость распределения различных видов гидроидов от характера и скорости течений.
Теперь настало время разобраться в том, какие особенности морфологии, биологии и экологии гидроидов определяют их зависимость от перемещения воды вокруг колонии; каково разнообразие экологических ниш, выделяемых по гидродинамическим критериям; какие морфогенетические механизмы лежат в основе приспособлений строения колоний у гидроидов к гидродинамическим условиям существования.
На эти вопросы пока нет исчерпывающих ответов. Однако подробное изучение морфологии колоний Obelia loveni [8], О. longissima [3] и О. geniculata дает возможность высказать ряд предположений о функциональной подоплеке различных особенностей строения гидроидов. Тем самым мы стремимся конкретизировать проблему, указать на возможные пути ее решения и привлечь специалистов по биомеханике и гидродинамике к разработке теории адаптации сессильных животных к гидродинамическим особенностям среды обитания.
В Великой Салме Ругозерской губы Белого моря, где мы .проводили наблюдения, О. geniculata обитает в местах с сильными приливоотливными течениями, в частности на Еремеевском пороге. Здесь нет прибойных волнений, а перемещение воды во время приливов и отливов имеет поступательный характер. Скорость течения все время плавно меняется и четыре раза в сутки достигает 70—80 см/с.
Ранее [5] было экспериментально доказано, что колонии О. geniculata хорошо растут в широком диапазоне скоростей обтекания их водой — от 9 до 51 см/с — и менее хорошо, но все же растут, при скорости 75 мс/с. Другие родственные виды, такие, как О. loveni и О. longissima, при такой скорости течения в эксперименте существовать уже не могут.
Теперь мы можем объяснить, каким образом некоторые особенности строения колонии О. geniculata помогают ей существовать на сильном течении. Прежде всего следует отметить, что побеги у О. geniculata по сравнению с другими видами обелий [3, 8] невысоки и в большинстве случаев слабо ветвятся, что уменьшает их парусность. Уплощенно-каплевидная форма междоузлий, свойственная только этому виду беломорских обелий, делает ствол более обтекаемым, улучшая тем самым его гидродинамические качества.
Большое значение для жизнедеятельности колоний имеет сопротивление побегов на изгиб. Например, у О. loveni сильное течение прижимает побеги к субстрату, а боковые веточки и гидротеки — к стволу побега [5]. В таком состоянии гидроид не может ловить добычу, а следовательно, и существовать в местах с сильным поступательным течением. У О. geniculata имеется ряд морфологических приспособлений, уменьшающих гибкость побегов. Прежде всего это отсутствие перетяжек на перисарке в основании каждого междоузлия. Регулярное расположение перетяжек в этой зоне делает перисарк как бы членистым, увеличивая его гибкость. Для представителей рода Obelia весьма характерно наличие хотя бы нескольких перетяжек в проксимальной части каждого междоузлия. Однако у О. geniculata перетяжки очень слабо выражены, членистости перисарка нет, что уменьшает гибкость побега. У других видов рода Obelia увеличение жесткости побега достигается другими путями — например, у О. longissima за счет утолщения перисарка и образования полисифонности [З].
Особое значение для сохранения побега в стоячем положении имеет жесткость его базальной части. Здесь, как и в каждом междоузлии побега обелий, должно быть несколько перетяжек. Однако у О. geniculata их нет. Кроме того, ножка побега у этого вида по сравнению с другими видами укороченная. Приведенные особенности морфологии можно рассматривать как приспособление к уменьшению гибкости побега без образования дополнительных структур или утолщения перисарка.
Одна из основных функций гидротек — защита гидрантов от механического воздействия течения, в котором тонкие щупальца не могут сохранять свое ловчее положение, так как прижимаются током воды друг к другу. Гидротеки служат экраном, своеобразными обтекателями для гидрантов. В потоке воды гидротеки обелий обращены к течению основаниями. Сами гидранты при этом не испытывают прямого действия течения, лишь их щупальца частично выставлены за пределы гидротек. У О. loveni гидротеки имеют форму конуса с узким основанием (устьем), и щупальца гидрантов поэтому почти целиком оказываются за их пределами. По сравнению с ними гидротеки О. geniculata имеют более широкое устье, и щупальца высовываются за его пределы лишь своими кончиками. Такая особенность формы гидротек способствует сохранению ловчего положения щупалец гидрантов О. ge-niculata на сильном течении.
Обладая относительно короткими ножками, широкие гидротеки О. geniculata оказываются менее гибкими, что, возможно, уменьшает их биение в потоке. Из-за малого угла наклона ножек к оси побега гидранты приближены к стволу. Небольшая асимметрия гидротек делает их еще более параллельными стволу побега, помогая при ограниченной гибкости ножек занять положение вдоль потока.
При жизни на сильных течениях колонии должны быть прочно прикреплены к субстрату. У О. geniculata это достигается за счет уплощения прилегающей к таллому водоросли стороны гидроризы, что обеспечивается своеобразной формой верхушек столонов — более широкой, чем у других обелий.
Образование фрустул у О. geniculata — не реакция гидроида на ухудшение условий существования, как предполагалось ранее [II], а одно из главных приспособлений к обитанию в местах с сильным течением, позволяющее увеличить размеры популяции там, где оседание личинок крайне затруднено. Фрустулы сразу прилипают к субстрату вблизи колонии. Часто этому способствует некоторый наклон побега к субстрату, во время которого фрустулообразующий отросток касается субстрата, прилипает к нему своей верхушкой, после чего происходит отделение его дистальной части, т. е. фрустулы. Таким образом, даже на сильном течении прилежащие территории могут быть освоены благодаря бесполому размножению. В начале лета это единственный способ увеличения численности популяции гидроида, так как половое размножение О. geniculata в районе наших исследований задерживается до июля.
Благодаря фрустуляции побеги у О. geniculata остаются слаборазветвленными, что уменьшает их парусность и, следовательно, вероятность отрыва сильным потоком воды. Важно отметить, что это достигается не столько сокращением ветвления побегов, сколько прерыванием развития молодых веточек, концы которых преобразуются во фрустулы.
Для экспериментальной проверки изложенных соображений относительно возможного функционального значения ряда морфологических признаков строения колонии О. geniculata мы провели специальное исследование состояния побегов и гидрантов нескольких видов гидроидов в потоке воды различной интенсивности. Непосредственные наблюдения за положением побегов и гидрантов О. geniculata в течении полностью подтвердили предположения о роли сглаженных перетяжек пересарка в увеличении жесткости побега, ограниченного ветвления и укорочения междоузлии в уменьшении парусности побегов, широких гидротек в ослаблении действия напора воды на щупальца.
Таким образом, отличительные морфологические признаки О. geniculata имеют важное функциональное значение, тесно связанное с адаптацией гидроида к существованию в местах с сильным течением.