Внутривидовая гибридизация хвойных пород
СОДЕРЖАНИЕ: Народнохозяйственное значение и селекция хвойных пород, их систематика и распространение, селекционные методы улучшения. Гибридизация хвойных пород, подходы к данному процессу и оценка полученных результатов. Принципы и этапы подбора родительских пар.Введение
Лесная селекция и генетика – наука об улучшении качества и повышении продуктивности лесов путем управления наследственностью и изменчивостью древесных и кустарниковых растений. Она неразрывно связана с лесным семеноводством, задача которого заключается в получении сортовых семян древесных растений и районировании их пользования.
В программу интенсификации всех отраслей растениеводства, в том числе и лесных культур, наряду с агроприемами, включается использование улучшенного селекционно-генетическими методами посевного и посадочного материала. Таким образом, сорт лесных древесных пород так же, как и сорт в сельском хозяйстве, становится средством производства. Сор том называют совокупность растений с улучшенными по сравнению с исходными видами и сортами хозяйственными и биологическими признаками, устойчиво передающимися потомству при семенном и вегетативном размножении. Селекцией называют процесс выведения новых – и улучшения существующих сортов растений и пород животных. Слово селекция в переводе с латинского означает отбор или выбор. Отбор лучших форм из дикорастущих видов растений – наиболее древний метод селекции. Первоначальное понятие селекции соответствовало содержанию работ по выведению новых сортов путем отбора и размножения лучших форм. Современная селекция применяет отбор, используя при создании исходного материала гибридизацию, полиплоидию и мутагенез. Однако и в настоящее время отбор остается основой селекционной работы: с отбора исходных форм начинается выведение сорта, отбором заканчивается апробация форм и признание нового сорта.
Наука, изучающая закономерности наследственности и изменчивости, называется генетикой. Наследственность неразрывно связана с изменчивостью. Закономерности наследственности и изменчивости растений, установленные и разрабатываемые генетикой, составляют основу селекционной работы по выведению новых сортов. Генетика же заимствует и использует для обобщения и установления закономерностей наследственности и изменчивости фактический материал, получаемый селекцией в процессе создания сортов. Селекция растений имеет и собственные задачи по управлению процессами сортоводства, которые являются одной из форм направленной эволюции. Селекция – это учение об искусственном отборе в широком смысле этого слова. Изучение закономерностей естественного отбора позволило Ч. Дарвину селекцию из ремесла и искусства превратить в науку. Несмотря на выделение генетики в самостоятельную науку, селекция, как учение о закономерностях выведения сортов на организменном и на популяционном уровне, не лишилась самостоятельности, а приобрела новый более совершенный уровень своего развития.
Селекционно-генетические и семеноводческие знания о лесных древесных и кустарниковых растениях входят в состав смежной науки – лесной селекции и генетики. Задача этой науки заключается в разработке селекционно-генетических методов повышения продуктивности и улучшения качества создаваемых лесов. Выполнение этой важнейшей задачи практической селекции в области лесоводства неотделимо от сортоводства древесных пород.
По способам размножения сорта древесных растений можно разделить на четыре группы: сорта-клоны, сорта-линии, сорта-популяции и сорта гибридного происхождения, или сорта-гибриды.
Сорта, получаемые скрещиванием особей с разной наследственностью и отбором из гибридных семей, называются гибридными. Эти сорта преобладают в настоящее время во всех областях растениеводства, так как они обладают выделяющимися качествами вследствие последовательного накопления на различных ступенях сложного скрещивания важнейших свойств, которыми должен обладать сорт.
В лесном хозяйстве и декоративном садоводстве использование диких видов и форм в культуре приобретает в настоящее время особое значение Возможность применения селекции к лесным древесным растениям не вызывает ни у кого сомнений, а переход при ведении лесного хозяйства от рода и вида к сорту – реальная практическая необходимость.
1. Селекция хвойных пород
1.1 Народнохозяйственное значение и селекция хвойных пород
Хвойные – основные породы-лесообразователи Советского Союза. Они занимают большие площади таежной зоны Евразии, составляя основу лесосырьевой базы страны. Хвойные леса СССР оказывают существенное влияние на климат Северного полушария планеты. Основное хозяйственное значение у хвойных имеет древесина, которая широко используется для изготовления пиломатериалов, в качестве сырья для целлюлозно-бумажной промышленности, для получения искусственного шелка, производства музыкальных инструментов, токарных изделий и т.д.
Древесина сосны с красноватым ядром и желтовато-белой заболонью с четкими границами годичных колец издавна пользуется большим спросом. Круглый лес (столбы, бревна, рудничная стойка), пиломатериалы (балки, брусья, доски, тес), колотый лес и щепные товары (клепка, щепа) – основные сортименты сосновой древесины. Мягкая ядровая древесина кедра сибирского используется для производства карандашей. Древесина лиственницы, очень стойкая в воде и противостоящая гниению, успешно используется для гидротехнических сооружений, подводных и мостовых брусьев, рудничной стойки и др. С красноватым ядром и желтовато-белой заболонью древесина лиственницы очень Декоративна, применяется дли отделки Интерьеров общественных зданий. Красоту рисунка древесины лиственницы дополняют четко выраженные годичные слои. Древесина богата смоляными ходами. Древесина ели и пихты без ядра. Она применяется как строевой лес, и как сырье для целлюлозно-бумажной промышленности, для получения искусственного шелка, и изготовления музыкальных инструментов. Древесина можжевельника отличается прочностью, не имеет смоляных ходов и обладает высокой декоративностью благодаря коричневатому ядру и узкой беловатой заболони. Годичные слои у нее извилистые, поздняя древесина их резко отличается от ранней. Древесина можжевельника пользуется большим спросом для изготовления токарных изделий, тростей, шахмат, игрушек и сувениров.
Для обширных областей Европы, Азии, Северной Америки и отчасти Африки древесина хвойных издавна была основным строительным материалом, из которого возводились жилища, хозяйственные постройки, культурные и общественные здания. Значение древесины хвойных как идеального по своим техническим качествам столярного и строительного материала особенно возросло за последние два столетия, когда механизация производства потребовала однородного сырья древесины. Однако наибольшее значение приобрела древесина хвойных в целлюлозно-бумажной промышленности, где ценится длина волокна. Трахеиды, из которых главным образом сложена древесина хвойных, оказались самым длинноволокнистым элементом среди современных древесных растений. Наибольшей длиной из хвойных северного полушария отличаются трахеиды ели (4300 мкм) и лжетсуги (4500 мкм).
Хвойные широко используются для добычи смолы, дубильных веществ и другого ценного сырья для промышленности. При подсочке хвойных добывают живицу для производства скипидара и канифоли, в коре ели и лиственницы содержатся дубильные вещества, из коры можжевельника добывают скипидар для лаков, а смола коры пихты содержит до 25% скипидара. Семена кедровых сосен и шишки можжевельников используются в питании человека. Из хвои пихты, ели, сосны и можжевельника получают эфирные масла и витамин С, используемые в медицине.
Продолжительная и интенсивная эксплуатация хвойных поставила человечество перед необходимостью забот о сохранении и селекции их на повышение продуктивности и качества древесины, живицы и др. Дальнейшая интенсификация лесного хозяйства предполагается не только за счет селекции местных хвойных видов, форм и сортов, но и широкой интродукции хозяйственно ценных зарубежных пород.
1.2 Систематика и распространение хвойных пород
Современные хвойные, или пиниды, насчитывают 5 порядков, 7 семейств, 57 родов и 560 видов.
Среди современных хвойных самыми древними семействами являются араукариевые, подокарповые и сосновые. Пыльцевой анализ показал, что предки современных представителей хвойных существовали еще до мелового периода, т.е. до появления цветковых растений. За время своего существования хвойные подверглись значительной эволюции. Однако хвойные отличаются заметной стабильностью генетической организации. Эволюционная стабильность хвойных объясняется общей консервативностью возникших в ходе эволюции хвойных морфологических структур. Говоря о темпах хромосомной эволюции, генетики отмечают, что у сосновых они в десятки и сотни раз ниже по сравнению с другими организмами. Вместе с тем скорость эволюции структурных генов, ответственных за синтез белков у сосновых, соответствует средней скорости эволюции других организмов.
Долговечность хвойных (от 300 до 2000–3000 лет) определяется особенностями строения стволов, отличающихся от стволов других голосеменных более развитой древесиной и менее развитыми кроной и сердцевиной. Ксилема хвойных по объему на 90–95% состоит из трахеид, длина которых у разных видов сильно варьирует (от 0,5 до 11 мм). В процессе эволюции длина трахеид постепенно уменьшается. Число пор на каждой трахеиде от 30 до 50 шт., они сосредоточены на концах трахеид, где интенсивность тока пасоки наибольшая. Трахеиды с округлыми окаймленными порами – достаточно совершенные водопроводящие элементы.
Древесной паренхимы у хвойных очень мало, у тисовых и араукариевых она отсутствует. Сердцевинные лучи узкие, однорядные или двурядные (реже более широкие) от 1 до 60 клеток по высоте.
В древесине и коре современных хвойных много смоляных ходов. Они имеют форму длинных узких каналов, наполненных эфирными маслами, смолой и бальзамами, выделяемыми выстилающими клетками. Только для представителей семейства сосновых характерны постоянные нормальные (не травматические) смоляные ходы во вторичной древесине. У огромного большинства хвойных, не имеющих смоляных ходов в древесине в норме, они образуются как реакция на поранение ствола. Однако у араукариевых смоляные ходы не образуются даже при повреждениях стволов. Смола у них откладывается в трахеидах, окаймляющих сердцевинные лучи.
Хвойные – разнополые, однодомные, реже двудомные растения. Микростробил, который часто называют мужским колоском, состоит из укороченной оси, на которой расположены сильно редуцированные микроспорофиллы с микроспорангиями. В систематики и филогении хвойных особый интерес представляет строение женских стробилов, которые называют макро- или мегастробилами. Уменьшение числа макростробил – общее направление эволюции хвойных. У тисовых в результате далеко зашедшей редукции числа макростробилов формируются одиночные, односеменные макростробилы. Эволюция большинства хвойных шла в сторону возрастания компактности собрания макростробилов, уменьшение же числа частей играло второстепенную роль. В итоге возникли компактные шишки – типа шишек сосны, ели, араукарии и других хвойных.
У большинства представителей семейства сосновых (сосны, ели, пихты) пыльца бобовидной формы с двумя выступами или воздушными мешками. Для видов тсуги типична шарообразная пыльца с одним опоясывающим воздушным мешком. У лиственницы и ложнотсуги пыльцевых мешков нет и пыльца имеет шарообразную форму.
Ветроопыление у хвойных, т.е. перенос пыльцы на семяпочку движением воздуха, происходит различно. Наиболее известный тип опыления характерен для сосны, имеющей пыльцу с двумя воздушными мешками. Семенные чешуйки в шишке сосны в период цветения несколько раздвигаются, что свидетельствует о готовности семяпочек к принятию пыльцы. Обильное количество пыльцы, доставляемое ветром, попадает в промежутки между семенными чешуями и задерживается так называемыми рыльцевыми расширениями края микропиле (пыльцевхода). У сосны эти расширения узкие, у ели они шире. К этому времени (а иногда и раньше) микропиле начинает выделять так называемую опылительную жидкость, выступающую на его верхушке в виде капли (опылительная капля). Пыльцевые зерна легко смачиваются опылительной жидкостью и быстро погружаются в нее. В результате пыльцевое зерно как бы засасывается внутрь и достигает верхушки мегаспорангия (нуцеллуса), где и прорастает. Благодаря воздушным мешкам, играющим роль своеобразных плавательных приспособлений, пыльцевое зерно по мере продвижения вверх по микропилярному каналу все время остается ориентированным дистальным полюсом вниз по направлению в пыльцевую камеру, образованную краями нуцеллуса. В таком положении пыльцевое зерно легко прорастает и образует пыльцевую трубку. После опыления чешуи сближаются и остаются плотно прижатыми друг к другу до созревания семян.
Развитие зародыша семени у хвойных начинается с образования предзародыша, или зачаточного зародыша (проэмбрио). После этого начинается развитие собственно зародыша. Увеличиваясь, подвесок не только проталкивает зародыш вглубь ткани гаметофита и служит для них гаусториальным органом, но и является, вероятно, секреторным органом, разрушающим с помощью ферментов клетки эндосперма.
Семена хвойных очень разнообразны по величине и форме. Крупные семена с деревянной кожурой бескрылые, а кожистые и перепончатые большинства хвойных снабжены одним крыловидным придатком или двумя-тремя небольшими крыльями, что способствует их распространению ветром. Сочный, ярко окрашенный покров на семенах ряда хвойных способствует их распространению животными.
Исчезновение воздушных мешков у пыльцы лиственницы и, лжетсуги связано с отсутствием опылительной жидкости. Следует, однако, отметить, что у пихты и кедра опылительная жидкость не выделяется, но пыльцевые зерна снабжены воздушыми мешками, хотя у кедра наблюдается тенденция к их редукции. У пихты микропилярная область превращается в косую рыльцевую воронку, улавливающую пыльцу. Пыльцевые зерна не достигают нуцеллуса. Вместо этого нуцеллус растет кверху, и этим достигается контакт с пыльцевыми зернами. У кедра, как и у пихты, в результате согласованных изменений микропиле и нуцеллуса пыльцевые зерна оказывая на нуцеллусе, где и прорастают.
Оплодотворение у хвойных, за исключением сосны, происходит вскоре после опыления. У видов сосны между опылением и оплодотворением проходит от 12 до 14 мес.
В процессе длительной эволюции ряд видов хвойных успешно приспособился к изменяющимся условиям среды благодаря совершенствованию и закреплению в потомстве древних генетических структур, определяющих соответственно проявление конкурентоспособных свойств. Подтверждением этого могут служить таежные леса с преобладанием хвойных пород, занимающие огромные территории холодной зоны в Северном полушарии, образующие чистые и смешанные хвойно-лиственные древостои. Ксероморфность ассимиляционного аппарата, Относительная долговечность, выработка защитных средств (живицы, эфирных масел), строение водопроводящей системы, ветроопыляемость и ряд других особенностей строения обеспечили хвойным одно из ведущих положений в образовании современных лесных формаций на земном шаре. Знание особенностей исторического развития, или филогении, видов позволяет выработать методы управления закономерностями наследственности и изменчивости при их искусственном разведении.
1.3 Селекционные методы улучшения хвойных пород
Селекционно-генетическое улучшение лесообразующих хвойных пород в нашей стране осуществляется в плановом порядке. Селекционные программы составляются по следующим направлениям: секция на быстроту роста и продуктивности сосны, ели, лиственницы, пихты, можжевельника; селекция на смолопродуктивность сосны; селекция на урожайность кедровых сосен; селекция на резонансность древесины ели; селекция на декоративность и устойчивость к промышленным выбросам сосны, ели, лиственницы, пихты, можжевельника, псевдотсуги, тсуги и др. Комплексное использование массового, семейного и клонового отборов – основной метод современной селекции хвойных.
Селекционная практика показала, что отбор целесообразно вести по небольшому числу признаков. Успех работы по селекции, в том числе и хвойных пород, во многом зависит от методики изучения прямых селекционных признаков и корреляции их с удобными для использования косвенными признаками. Генетическая обусловленность признака у лесообразующих хвойных определяется путем математического разложения общей фенотипической изменчивости на генотипическую и паратипическую (средовую) изменчивость. Наследуемость в общем смысле, без смены поколений в природных популяциях хвойных, изучается дисперсионным анализом и другими математическими методами при отборе на общую комбинативную способность. Для определения наследуемости в узком смысле слова требуется изучение характера наследования признака (доминирование, эпистаз или аддитивное наследование), которое используется при отборе на специфическую комбинативную способность. Эти знания необходимы для практической работы по селекции, так как они позволяют раскрыть генетическую характеристику селекционного признака.
Селекционно-генетическая программа улучшения хвойных включает такие вопросы:
1. Постановка цели и задачи. Современное состояние вопроса и перспективы на обозримое будущее (5, 10–20, 50 и более лет).
2. Исходный материал для селекции (родовой комплекс лесообразователей местных пород и интродуцентов).
3. Методы отбора и интенсивность ведения хозяйства (массовый и групповой отбор с определением общей комбинативной способности в диких популяциях.
4. Отбор на общую и специфическую комбинативную способность, индивидуальный отбор и плантационное хозяйство.
5. Создание постоянной семеноводческой базы для получения сортовых семян в соответствии с направлением ведения хозяйства.
6. Организация выращивания сортового посевного и посадочного материала для лесных культур и промышленных плантаций.
7. Закладка культур и промышленных плантаций по выращиванию заданных сортиментов древесины.
8. Разработка селекционных методов рубок ухода, стимулирующих рост деревьев в искусственных посадках.
Таким образом, селекционные методы улучшения хвойных органически входят в общую систему лесоводческих мероприятий по интенсификации лесохозяйственного производства. Особое значение в деле повышения производительности хвойных лесов нашей страны приобретает создание популяционного сортоводства на основе системного генетического анализа естественных насаждений. Поэтому знания частной генетики хвойных в целом, а также отдельных родовых комплексов и видов необходимы лесоводам, чтобы успешно осуществлять селекционно-семеноводческую программу в лесах нашей страны.
1.4 Исходный материал для селекции
Селекционная работа начинается с подбора исходного материала в соответствии с избранным направлением и поставленной целью. Исходным материалом в селекции называют культурные и дикие формы растений и популяций, используемые для выведения новых сортов. В современной селекции в качестве исходного материала используются несколько форм и популяций.
Естественные (природные) популяции – дикорастущие формы, местные сорта культурных растений и образцы мировой коллекции сельскохозяйственных растений Всесоюзного института растениеводства имени Н.И. Вавилова.
Гибридные популяции – внутривидовые популяции, создаваемые в результате скрещивания сортов и форм в пределах одного вида, и популяции, полученные от скрещивания особей разных видов и родов растений (межвидовые и межродовые).
У перекрестноопыляющихся растений одним из новых источников материала являются самоопыляемые линии, или инцухт-линии. Их получают многократным принудительным самоопылением перекрестноопыляющихся растений. Лучшие линии скрещивают между собой, получают межлинейные гибриды или их скрещивают с другими сортами для создания гетерозисных гибридов. В результате скрещивания получают гибридные семена, которые используют в течение 1 года. Сорта-гибриды этих межлинейных скрещиваний в отличие от сортов гибридного происхождения нужно ежегодно воспроизводить.
Искусственные мутации и полиплоидные формы создаются путем воздействия на растения различными мутагенами (радиацией, химическими веществами, температурой, прививками и др.).
Ценным источником сортов древесных растений являются формовые разнообразия естественных популяций и интродукции новых видов из других флористических зон. Введение в культуру новых форм и сортов в будущем улучшит качество наших лесов. Селекция поможет создавать сортовые промышленные плантации в соответствии с ростом потребности в определенных сортиментах древесины и других полезностях леса.
В процессе развития человечества наши леса, как и остальные компоненты природы, подвергаются значительным изменениям. Так, в течение многих столетии выборочные рубки деревьев лучшего качества без заботы о восстановлении лесов отрицательно повлияли на лесонасаждения. Это сокращает мировой лесной генофонд лесных древесных популяций.
Работа по селекции всех видов растений, в том числе и древесных пород, основывается на использовании естественного генофонда местных древесных пород лесообразователей и интродуцентов. Сохранение генофонда древесных растений как для осуществления мероприятий по улучшению свойств древесных растении в настоящее время, так и для селекционной работы и составлении перспективного ассортимента в будущем становится в центре внимания лесной селекции. Учет и исследование генофонда – основа каждой селекционно-генетической программы.
Все особи вида имеют общие характерные черты, а также свои индивидуальные генотипические признаки, что в целом отражает генетическое разнообразие вида, которое называют генофондом вида.
Генофонд древесных растений обследуется в настоящее время путем изучения генетической структуры популяций основных лесообразователей. При выборе признаков, фенотипическое проявление которых могло бы дать информацию о генофонде популяции, большое значение имеют два показателя: генетический и экономический. Генетическая характеристика генофонда включает степень наследуемости признаков, выражающую сущность фенотипической ценности популяции. Анализ генофонда популяции возможен именно по фенотипическим выражениям тех признаков, которые характеризуются высокой степенью наследуемости. В свою очередь степень наследуемости определяется по регрессии «родители – потомки», анализу сибсов и полусибсов и в некоторых случаях проверкой коэффициента наследуемости аналогичной изменчивости эталонным методом в географических клоновых культурах с выявлением соотношения между генетической и фенотипической дисперсиями.
Экономическая характеристика генофонда имеет важное значение при изучении генофонда по хозяйственно ценным признакам. Но при этом не следует забывать, что пока еще невозможно уверенно прогнозировать экономическую ценность многих признаков для будущего. Это во многом зависит от уровня развития промышленности по использованию и переработке древесины и других полезностей лесных древесных растений в будущем. В связи с этим необходима разработка мероприятий по сохранению в наших лесах всей полноты генофонда. Поэтому селекционные размеры популяций должны быть большими, что возможно при сохранении значительного числа особей каждой популяции.
2. Гибридизация хвойных пород
2.1 Гибридизация
Скрещивание особей, отличающихся друг от друга хотя бы одним аллелем, называется гибридизацией. Гибридами соответственно называют потомство, полученное в результате искусственного или свободного (спонтанного) скрещивания особей с разной наследственностью. Различают внутривидовую, межродовую и межсемейственную гибридизацию. Скрещивание особей различных форм и сортов, принадлежащих к одному виду, называется внутривидовой гибридизацией. Скрещивание особей, принадлежащих к разным видам одного рода, разным родам и разным семействам, называется отдаленной межвидовой, межродовой и межсемейственной гибридизацией.
Гибридизацию нельзя рассматривать как простое арифметическое суммирование признаков и свойств растений. Родительские организмы передают потомству не признаки, а гены, на основе которых в каждом поколении гибридов признаки, контролируемые этими генами, развиваются вновь. Гибридизация используется в качестве способа изучения наследования, получившего название гибридологического метода генетического анализа. Этот метод генетики основан на принципе менделеевского анализа наследования и взаимодействия отдельных генов у организмов. При этом у гибридного потомства изучается наследование не совокупности признаков, а одного, двух или трех контрастных признаков в ряду последовательных поколений с применением индивидуального анализа потомства от каждого гибридного растения.
2.1.1 Гибридизация как метод селекции
Гибридизация как метод селекции включает комплекс приемов, направленных на получение гибридных растений с изменением наследственности и использованием ее для выведения новых сортов. Создавая гибридизацией нужный исходный материал, удается значительно ускорить ход селекционного процесса. Последовательным скрещиванием наследственно расщепляющихся родительских форм селекционеры создают новые формы растений. Выведение новых сортов, в которых с помощью гибридизации достигается сочетание хозяйственно ценных свойств большого количества родительских форм названо синтетической селекцией. Гибридизацию относят к категории комбинативной селекции, так как основной целью при этом является получение потомства с новой совокупностью генетически обусловленных признаков и свойств. Последующим отбором и направленным воспитанием гибридного потомства новые ценные признаки и свойства закрепляются и усиливаются.
2.1.2 Содержание метода гибридизации
Содержание и порядок работы по селекции методом гибридизации:
1. Цель работы и разработка модели (образа) будущего гибридного сорта.
2. Изучение генетического потенциала (наследственности) исходного материала.
3. Подбор родительских пар.
4. Подбор и хранение пыльцы.
5. Подготовка женских цветков к опылению (кастрация и изоляция).
6. Проведение опыления (техника скрещивания).
7. Наблюдение за развитием гибридных семян и уход за материнскими растениями.
8. Сбор гибридных семян и выращивание гибридного потомства.
9. Отбор лучших гибридных форм и выделение из них отдельных растений (кандидатов в сорта) для сравнительного испытания на сортоиспытательных участках.
10. Разработка методов массового размножения нового сорта для производства.
Сортам, полученным в итоге гибридизации, присваивается название, которое включает название исходных сортов или видов. Очень часто селекционеры присваивают сортам произвольные названия, подчеркивая их характерные сортовые особенности. Например, лесными селекционерами СССР получены региональные зимостойкие сорта белого пирамидального тополя от скрещивания местных форм белого тополя с тополем Боллеана. Этим сортам даны соответствующие названия: Советский пирамидальный селекции А.С. Яблокова, Уральский пирамидальный селекции Н.Н. Коновалова и т.д.
2.2 Типы скрещиваний, применяемые при гибридизации
Изменение наследственности исходного материала для селекции при гибризации осуществляется однократными, или простыми, скрещиваниями и многократными скрещиваниями, названными сложными или ступенчатыми скрещиваниями. Наиболее распространенное деление скрещиваний на простые и сложные с последующим подразделением на целый ряд типов можно представить такой схемой.
Простыми скрещиваниями называют однократные скрещивания между двумя родительскими формами. Если родительские виды или сорта обозначить буквами, то этот тип скрещивания можно изобразить как АЧБ или ВЧГ и т, д., после которых в гибридном потомстве проводится отбор элитных растений и оценка их потомства. При простых скрещиваниях гибриды получаются на основе комбинаций генов материнской и отцовской форм. Простые парные скрещивания имеют большее значение при внутривидовой гибридизации.
Диаллельные скрещивания – каждая испытываемая линия, форма или сорт скрещивается со всеми другими линиями или сортами во всех возможных комбинациях. Например, А ЧБ , АЧВ , АЧГ , АЧД , АЧЕ и т.д. Число всех возможных комбинаций при диаллельных скрещиваниях может быть очень большим и будет возрастать – по мере увеличения количества исходных линий, форм или сортов. Так, при диаллельном скрещивании 10 линий число всех возможных комбинаций составляет 45, а при 100 линиях уже 4950. Поэтому на практике диаллельные скрещивания обычно проводят в пределах сравнительно небольших групп, в которых бывает примерно по 10 линий или сортов. Диаллельные скрещивания можно применять в работе по гибридизации с древесными растениями с целью изучения варьирования признаков в гибридном потомстве, а также определения отобранных по фенотипу деревьев на проявление хозяйственно ценного признака в гибридном потомстве.
![]()
![]()
![]()
![]()
![]()

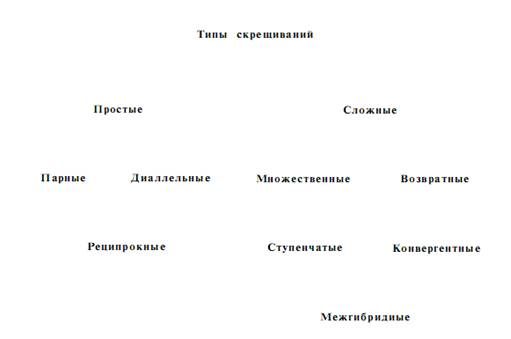
Реципрокными скрещиваниями называются скрещивания растений, при которых каждый из двух сортов или видов в одном случае является материнской формой, во втором – отцовской. Например, скрещивание, осуществленное по схеме АЧБ и БЧА . Первое скрещивание АЧБ называют прямым, второе БЧА –обратным. Реципрокные скрещивания часто используют с разведывательной целью, чтобы выяснить какую из двух форм лучше взять в качестве материнской, а какую в качестве отцовской. Реципрокные скрещивания важны при отдаленной гибридизации, так как часто успех работы решается удачным подбором отцовских и материнских видов. Для реципрокных скрещиваний составляют план, или сетку, скрещиваний.
Сложными скрещиваниями называются скрещивания, когда в гибридизацию вовлекается более двух родительских форм или когда гибридное потомство повторно скрещивается с одним из родителей. Сложные скрещивания в селекционной практике имеют значительно большее значение, чем простые.
Множественные скрещивания, или поликросы, – это такие скрещивания, когда материнское растение опыляется смесью пыльцы нескольких видов и сортов. Этот вид скрещивания схематически можно изобразить так: АЧ (Б+В+Г+Д и т.д.), где буквой А обозначен материнский сорт, а буквами Б , В , Г , Д – отцовские сорта, от которых берется пыльца для составления смеси. Множественные скрещивания осуществляются двумя способами: искусственным опылением материнского растения смесью пыльцы нескольких отцовских форм: свободным опылением материнского растения с помощью ветра или насекомых, когда материнские и отцовские растения высаживаются рядом на одной и той же семенной площадке. Метод множественного опыления успешно применил к плодовым культурам И.В. Мичурин. В селекции многих кормовых многолетних трав и некоторых древесных растений сейчас широко используется этот метод, получивший название метода поликроссной селекции.
Основные условия успешного применения метода множественных скрещиваний: растения должны быть многолетними, самостерильными, обладать способностью к клонированию и иметь одинаковые сроки цветения. Таким условиям могут удовлетворять многие древесные породы.
Возвратные скрещивания, или беккроссы, – скрещивания, при которых гибрид повторно скрещивается с одной из родительских форм. В природных условиях повторные скрещивания спонтанных гибридов с одной из родительских форм называются интрогрессивной гибридизацией. В практике селекционной работы беккроссы называют повторными (возвратными) скрещиваниями. Возвратные скрещивания можно записать схематически так: 1-й год – А ЧБ ; 2-й год – (АЧ Б ) ЧА или (АЧБ )ЧБ . Этот тип скрещивания широко применяется в селекционной практике. Он используется в тех случаях, когда у ценных по комплексу признаков сортов имеется дефект, который желательно устранить. Тогда новый сорт стал бы совершеннее, расширились бы возможности его практического использования. При возвратных скрещиваниях тот сорт, от которого хотят взять основной комплекс признаков, берется при первом скрещивании обычно в качестве материнского, а при повторных скрещиваниях он используется в качестве отцовского. Изменчивость гибридного потомства при возвратных скрещиваниях суживается. Полученные гибриды дают расщепление в отношении 1:1 (1-й год – ааЧАА –Аа ; 2-й год – АаЧаа-1АаЧ1 аа ). Селекционная цель достигается сравнительно быстро.
Насыщающие и конвергентные скрещивания – повторные возвратные скрещивания. Этот метод часто применяется при выведении сортов устойчивых к болезням. При насыщающих скрещиваниях признаки и свойства одного из родителей почти полностью вытесняются за исключением немногих генов. Во многих случаях это бывает нежелательным. Во избежание этого разработана система конвергентных скрещиваний. Конвергентные скрещивания представляют собой дальнейшее развитие метода возвратных скрещиваний. Метод заключается в том, что после получения F 1 дальнейшее скрещивание проводят в двух направлениях. В одном случае гибриды повторно скрещивают с материнским сортом, а во втором – с отцовским. В результате получают две сближенные линии. Их скрещивают между собой и среди гибридного потомства производят отбор. После возвратных скрещиваний и сближения линий гибридное потомство проявляет менее сложный характер расщепления. Вследствие этого среди потомства легче найти желаемую комбинацию признаков.
Ступенчатые скрещивания – полученный от простого скрещивания гибрид повторно скрещивается не с родительской формой, а с третьим сортом или видом растений, затем с четвертым и т.д. Таким образом, в этих скрещиваниях участвуют несколько родительских форм, которые последовательно или ступенчато включаются в гибридизацию. При ступенчатых скрещиваниях создается гибридный материал, включающий наследственные свойства нескольких сортов, или видов растений.
Межгибридными скрещиваниями называют такие, при которых объединение наследственности нескольких родителей осуществляют не последовательно, как при ступенчатой гибридизации, а параллельно после предварительного получения простых гибридов и последующего их скрещивания.
Межгибридные скрещивания–главный метод при создании гетерозисных гибридных семян кукурузы и некоторых других культур.
Все типы скрещиваний успешно применяются к лесным древесным растениям. Они позволяют в ряде случаев создавать оригинальные комплексные программы применительно к особенностям той или иной группы видов. Перспективным приемом создания таких программ является включение внутривидовой и межвидовой гибридизации по родовым комплексам.
Лесные древесные растения выращиваются обычно с целью получения вегетативной массы, поэтому стерильность гибридных растений не может препятствовать их выращиванию в промышленных масштабах. В современной лесной генетике и селекции гибридизация играет более важную роль, чем в селекции сельскохозяйственных культур. Применение межвидовой гибридизации близких видов в некоторых родовых комплексах древесных пород из разных районов произрастания имеет большое значение для получения гетерозисных межвидовых гибридов. Если такие географически изолированные виды отобрать и посадить вместе, то можно получить гибрид, между ними от свободного переопыления.
Накоплены многочисленные экспериментальные данные, доказывающие целесообразность широкого применения внутривидовой гибридизации к лесным древесным растениям Проявление гетерозиса у лесных пород чаще наблюдается в первом гибридном поколении. Однако в некоторых случаях рациональнее селекционную работу распространять на второе и третье поколения. В связи с длительным периодом смены поколений у лесных древесных растении большое значение приобретает умелый подбор родительских пар с целью обеспечения в первом поколении гибрида с желательной комбинацией хозяйственно ценных признаков.
2.3 Принципы подбора родительских пар
хвойный гибридизация порода родительский
Общим показателем при подборе родительских пар для гибридизации является высокая выраженность у них интересующих селекционера признаков и свойств. Для правильного выбора пар требуется проверка наследования отдельных хозяйственно ценных признаков и свойств. К искусственно создаваемым гибридным популяциям лесных древесных пород предъявляются определенные требования: они должны обладать не только экономически выгодной комбинацией признаков, но и экологической и возрастной стабильностью их проявления.
Подбор пар по экологическому признаку был успешно применен И.В. Мичуриным. Учет исторически сложившихся биологических требований родительских организмов, изучение условий существования их предков позволили ему предвидеть возможный ход развития молодого гибридного растения в новых условиях. Он обнаружил, что при скрещивании местных сортов с инорайонными, первые, как более приспособленные к данным условиям, в большей степени передают свои признаки и свойства потомству.
Подбор пар по продолжительности отдельных фаз вегетации применяется при работе с целым рядом культур. Это позволяет проводить селекцию на скороспелость. В мировой селекционной практике отмечены случаи, когда гибриды или их потомство превосходят обоих родителей по тем или иным признакам и свойствам. Эти сдвиги могут быть как в сторону увеличения, так и в сторону уменьшения значения признака. Шведский селекционер Нильсон-Эле это явление назвал трансгрессией.
В результате исследований установлено, что трансгрессия проявляется в отношении таких признаков и свойств, которые контролируются не одним геном, а несколькими распределенными между скрещиваемыми партнерами. По мере увеличения у одной особи количества этих генов соответствующий признак проявляется сильнее или, наоборот, ослабляется. При скрещивании двух сортов, у которых тот или иной признак или свойство являются полимерными, у гибридного потомства возможно суммирование соответствующих генов и благодаря их однозначному действию произойдет усиление или, ослабление проявления данного признака или свойства.
Направление в селекции, основанное на планомерном ^скрещивании с целью получения положительных трансгрессий, называют трансгрессивной селекцией. Следует отливать трансгрессию от гетерозиса. При гетерозисе, в отличие от трансгрессии, гибридные растения имеют максимальную высоту всегда в первом поколении F. В последующих генерациях наблюдается быстрое снижение роста. Поэтому при семенном размножении растений невозможно закрепить гетерозис. У древесных растений гетерозисные формы размножают вегетативным способом. Если быстрый рост вызван трансгрессией, то это изменение наследуется во всех последующих поколениях, несмотря на расщепление гетерозигот.
В селекции кукурузы, сорго и других видов при подборе родительских пар широко используются электронно-вычислительные машины, особенно при обширных коллекциях сортов и форм. На перфорационные карты заносятся все данные, характеризующие имеющиеся в коллекции образцы по всем интересующим селекционера признакам и свойствам. На каждый образец заполняется отдельная карточка. Машине дается задание – решить при скрещивании от каких сортов, форм и линий можно с большей вероятностью получить наилучшее сочетание признаков и свойств у гибридов. Однако следует помнить, что правильность ответа определяется, прежде всего, тем, насколько полная и точная характеристика дается исходному материалу. Для использования ЭВМ необходимо предварительное детальное изучение исходных форм.
2.4 Способы получения гибридных семян древесных растений
Гибридные семена древесных растений получают от скрещивания на растущих корнесобственных и привитых деревьях, а также на срезанных ветвях. В крону деревьев для работы по искусственному опылению поднимаются при помощи лестниц, лазов, специально построенных подмостков и автомашин с подъемными приспособлениями.
Подготовка материнских деревьев для опыления у ветроопыляемых разнополых растений резко отличается от подготовки насекомоопыляемых обоеполых. У первых изоляцию проводят удалением мужских соцветий, (например, сережек у березовых и. стробилов у хвойных). У двудомных возможна пространственная изоляция. У обоеполых растений для изоляции женских цветков необходимо удалить из цветка пыльники, не повредив пестики. Перед изоляцией удаляют пинцетом или ножницами пыльники. Женские цветки и соцветия изолируют за несколько дней до начала цветения (пыления), чтобы предотвратить опыление их нежелательной пыльцой. Изоляторы изготовляют в виде пакетов или небольших мешочков из плотной ткани или бумажной кальки. Для изоляции цветков насекомоопыляемых растений используются марлевые мешочки. На основание изолируемой ветки, где изолятор завязывают шпагатом, подкладывают кусочек ваты для доступа воздуха в изолятор.
Пыльцу с 1 дерева-опылителя заготавливают заранее. У ветроопыляемых растений, мужские цветки которых собраны в соцветия (у буковых, березовых, тополей, ореховых) и в стробилы (у хвойных) для сбора пыльцы используют соцветия, которые снимают с растущих деревьев за 1–2 дня доестественного созревания (пыления). Собранные сережки или стробилы для дозревания помещают на бумажную кальку в один слой, где пыльники слегка подсыхают. Пыльцу собирают в пакеты или небольшие стеклянные емкости. Для сбора пыльцы в большом количестве целесообразно использовать почвенные сита, в которых пыльцу удобно очищать и вымолачивать из пыльников легким встряхиванием системы ситец. Так можно собирать пыльцу для массового опыления и получения гибридных семян от контролируемого опыления. Для сбора пыльцы из обоеполых цветков насекомоопыляемых растений тычинки выщипывают перед созреванием пыльников. Пыльники тонким слоем помещают в стеклянную посуду.
Если цветки на материнском дереве распускаются раньше, то для ускорения созревания мужских цветков прибегают к выгонке пыльцы. Для этого заготовляют ветки с цветками опылителя незадолго до цветения и помещают их в банку с водой в теплом месте. Банку ставят на гладкую бумагу, чтобы на нее осыпалась созревшая пыльца, которую собирают в стеклянные баночки или пакетики из бумажной кальки. Паспорт пыльцы составляют в дневнике по гибридизации, где указывают дату заготовки, номер дерева, с которого она собрана, и описание дерева. На этикетке можно написать номер опылителя и дату сбора. Для скрещивания географически отдаленных растений или видов с разными сроками цветения необходимо хранить и пересылать пыльцу по почте. Пыльцу хранят в эксикаторах с хлористым кальцием при температуре не выше 2–5°С (в холодильнике).
Продолжительность хранения пыльцы зависит от способа хранения и наследственных особенностей вида. Установлено, что пыльца березы, тополя, ивы и других видов может храниться (без применения) не более 1 мес., у сосны – свыше 1 года. Жизнеспособность пыльцы дуба при температуре 0°С и относительной влажности около 60% сохраняется более 2 мес. При низкой влажности пыльца дуба быстро погибает. Перед скрещиванием пыльцу, особенно долго хранившуюся или перенесшую заморозки, проверяют на жизнеспособность. О жизнеспособности пыльцы судят по проценту проросших пыльцевых зерен и длине пыльцевых трубок, развивающихся на питательной среде. Пыльцу проращивают в висячей капле или посевом на агар-агаровую поверхность в чашке Петри. В висячей капле пыльца сосны прорастает через 12–24 ч, а пыльца березы через 2–3 ч, что можно наблюдать под микроскопом.
Для опыления лучше пользоваться пыльцой, прорастающей на 50% и более. Искусственное опыление проводят в период, когда рыльца пестиков находятся в стадии оптимальной влажности и готовности к прорастанию пыльцы. У некоторых растений это состояние определяется выделением секрета в виде небольших капелек на рыльце пестика у покрытосеменных или в пазухах семенных чешуи у голосеменных растений. Женские цветки опыляют при помощи мягкой акварельной кисточки (рис. 56), если пыльцы немного, или создают пыльцевое облако не снимая изолятора с ветки. Облако пыльцы можно выпустить из пульверизатора или выдуть из стеклянной пипетки. В работах, где требуется абсолютная изоляция для опыления сосны, лиственницы, ели, березы и других пород при наличии большого количества пыльцы опыление производят при помощи шприца. Пакет прокалывают, вдувают в него пыльцу, затем проколотое отверстие заклеивают.
В северных районах опыление следует проводить в полдень, на юге – утром, при влажном воздухе. Опыление повторяют в течение 3 дней до утраты восприимчивости рылец. В северных районах рекомендуется материнские растения заключать в полиэтиленовые теплицы, где температурными воздействиями и изменением состава воздуха можно добиться раннего формирования обильного количества женских сережек на материнских деревьях. Пространственная изоляция этих деревьев упрощает работу по контролируемому опылению. После опыления на ветку необходимо повесить этикетку с шифрованным номером. В дневнике под этим номером записываются все сведения о родительских деревьях, особенно о проведении опыления. Полезно в шифровке указывать год работы, тогда по номерам будет легче определить время получения гибридных семян.
2.5 Скрещивание на срезанных ветвях
При гибридизации древесных растений с мелкими плодами и семенами (тополь, ива, ильмовые, береза) широко используется метод скрещивания на срезанных ветвях. В СССР и за рубежом гибридизацию ивовых проводят на срезанных ветвях. Для этого за 1,5–2 мес. до начала цветения срезают ветки длиной 1–1,5 м, диаметром от 0,6 до 2 см, с цветочными почками, на которых оставляют не более 10–12 цветочных и 5 листовых почек. Ветви помещают в колодезную или речную воду, которую меняют через 6–8 дней до начала цветения и через 3–5 дней с начала цветения и до созревания плодов. Срезы подновляют. В длительных опытах используются питательные растворы.
Ветки с мужскими цветками заготовляют в несколько сроков, но раньше, чем с женскими, чтобы, по возможности, совместить сроки цветения. Для пространственной изоляции ветви с тычиночными и пестичными сережками держат в разных помещениях. Техника опыления на срезанных ветвях такая же как и на растущих деревьях, но проводится оно на месяц раньше естественного цветения и позволяет получить гибридные семена к весеннему сроку посева.
Гибридизация на срезанных ветвях, кроме достоинств по технике скрещивания, позволяет проводить направленный уход за развитием гибридных семян. При скрещиваний на срезанных ветвях этих же видов тополей удалось получить около 50% зимостойких гибридов с пирамидальной кроной тополя Болле. Однако ослабление материнского растения допускается до определенного предела. Рекомендуется использовать для скрещивания тополей на срезанных ветвях длинные (0,6–1,5 м) и толстые (1,5–2 см) ветви с большим запасом питательных веществ. Среди полученных гибридных сеянцев (41 тыс.) на крупных ветвях ему удалось отобрать значительное число гетерозисных растений.
2.6 Способы выращивания и испытания гибридных растений древесных пород
Выращивать растения из гибридных семян необходимо на высоком агрофоне, чтобы обеспечить высокую грунтовую всхожесть и максимальную сохранность всходов, сеянцев и саженцев. Так, при получении гибридных семян осины, тополя и ивы гибридные всходы выращивают в чашках Петри, помещенных в термостаты, чтобы сохранить все гибридные растения и проследить за наследованием и характером расщепления хозяйственно ценных признаков и свойств. Выращивание березы на высоком уровне сохранности всходов показывает, что из одних и тех же гибридных вариантов можно вырастить от 2 до 300 гибридных всходов из одной сережки (в сережке березы бородавчатой содержится 500–600 семян). В зависимости от целей и направления селекции сеянцы и саженцы следует выращивать по-разному. При селекции на устойчивость гибриды надо выращивать на так называемом провокационном фоне, чтобы вести отбор на разных стадиях онтогенеза.
Выращивать гибридные растения следует с одновременным испытанием гибридного потомства. Подбор площадей под посев, выбор схемы посевов, методы посева и наблюдения за появлением всходов, ростом и развитием гибридных растений, оценка, отбор и выбраковка гибридных семян и отдельных растений должны быть подчинены конечной цели работы и проводиться при постоянном сравнении с контролем. Прежде всего необходимо сравнивать поведение растений, выращенных из семян от свободного опыления, с вариантами искусственного опыления. В связи с этим опыты по гибридизации следует ставить в нескольких повторностях и гибридное потомство выращивать также в достаточных повторностях. Некоторые селекционеры рекомендуют проводить все в четырех повторностях и более, однако во всех случаях следует рассчитать достоверность опыта.
Успешные примеры межвидовой и межродовой гибридизаций лесных древесных растений показали перспективность селекции путем гибридизации. В СССР и в зарубежных странах межвидовая гибридизация с отбором лучших тополей для вегетативного размножения на промышленных плантациях используется в лесоразведении при ускоренном выращивании сырья для целлюлозно-бумажной промышленности. Не меньшее значение в интенсификации лесного хозяйства имеет селекция на быстроту роста хвойных.
Таким образом, испытание потомства в гибридных популяциях необходимо вести по частной методике применительно к биологии данной породы и в соответствии с задачами, поставленными перед селекционерами промышленностью.
2.7 Гетерозис
Эффект гибридной мощности, или гетерозиса, заключается в превосходстве гибридов первого поколения по ряду признаков и свойств по сравнению с исходными родительскими организмами. Скрещивание гибридов между собой снижает этот эффект в следующих поколениях. Ч. Дарвин, попытавшийся объяснить природу этого явления, считал гетерозис одной из причин полезности скрещивания в эволюции видов. Перекрестное оплодотворение поддерживается естественным отбором именно потому, что оно служит механизмом поддержания гибридной мощности.
Эффект гетерозиса с позиций основных генетических закономерностей открытых в начале XX в. объясняется:
1) гетерозиготным состоянием гибридов по многим генам;
2) возникновением взаимодействия доминантных благоприятных генов;
3) сверхдоминированием – гетерозигота превосходит гомозиготу.
В доказательство первого положения приводятся примеры получения межлинейных гибридов. При скрещивании гомозиготных инбредных линий гибриды первого поколения гетерозиготны по многим генам. При этом действие вредных рецессивных мутантных аллелей подавляется доминантными аллелями обоих родителей. Например, одна инбредная линия в гомозиготном состоянии имеет рецессивную аллель одного гена ааББ , а вторая–другого гена ААББ . Каждая из этих рецессивных аллелей в гомозиготном состоянии определяет какую-нибудь недостаточность, которая снижает жизнеспособность инбредной линии. При скрещивании линий ааББ хААББ у гибридов объединяются доминантные аллели обоих генов – АаБб . Гибриды F 1 проявят при этом не только гетерозис по указанным генам, но и единообразие. В F 2 число особей с двумя доминантными генами в гетерозиготном состоянии АаБб будет лишь 4:1, поэтому гетерозисными оказываются не все особи. В дальнейших поколениях число гетерозигот сокращается, а число гомозигот увеличивается. Вследствие этого гетерозис в последующих поколениях затухает.
Однако не всегда гетерозиготность связана с гетерозисом. По второй гипотезе причиной гетерозиса является благоприятное действие некоторых доминантных аллелей. Поэтому подбор в гибридной комбинации доминантных аллелей может скорее обеспечить гетерозис. Таким образом, эта гипотеза исходит из представления о простом суммировании эффекта доминантных аллелей с комплементарным действием.
Третья гипотеза как бы развивает вторую. Она исходит из того, что гетерозиготное состояние аллелей имеет превосходство над гомозиготным. Следовательно, можно предположить, что сочетание в гетерозиготе аллелей дикого типа и мутантного каким-то образом усиливает действие доминантного гена и в связи с этим вызывает максимальное накопление специфических веществ, синтез которых контролируется этим геном. Указанное объяснение гетерозиса называют гипотезой сверхдоминирования.
Гипотеза доминантности удовлетворительно объясняет то ухудшение, которое наблюдается в результате инбридинга, а также восстановление мощности роста при скрещивании. На основании этой теории трудно, однако, объяснить, почему, гибриды по мощности превосходят популяции, из которых были получены их родители. Это увеличение мощности, по-видимому, следует объяснить сверхдоминантностью. При этом очевидно, что только небольшая часть локусов должна содержать гены, проявляющие сверхдоминантность в гетерозиготном состоянии.
Таким образом, две основные гипотезы, объясняющие гетерозис, ведут в общем к одинаковому результату и поэтому не являются взаимоисключающими. Имеются основания полагать, что гетерозис – более сложное явление и не может обусловливаться только одним типом взаимодействия.
Большой хозяйственный интерес представляют пути закрепления гетерозиса у древесных растений. Сохранение эффекта гетерозиса осуществляется вегетативным размножением уникальных гибридов.
2.8 Причины стерильности гибридов
Можно принять за правило, что чем дальше стоят друг от друга скрещиваемые виды и роды, тем сильнее выражена стерильность их гибридов.
Среди отдаленных скрещиваний различают две группы:
1) конгруентные скрещивания, когда родительские формы, несмотря на различие в генах, могут скрещиваться без понижения жизнеспособности, и
2) инконгруентные скрещивания, когда родительские формы имеют несоответственные хромосомы или разное число хромосом или несоответствие в плазме; у гибридов от таких скрещиваний обычно наблюдается неправильный мейоз или ненормальность в развитии и они оказываются стерильными.
Факторы, вызывающие стерильность, весьма различны:
1) Несовместимость ядра и цитоплазмы и как следствие этого – нарушение митозов в процессе развития генеративных тканей;
2) Действие генов, препятствующих развитию женских и мужских органов цветка;
3) Генетические факторы, препятствующие конъюгации хромосом в мейозе и образованию бивалентов, следствием чего является образование гамет с несбалансированным набором хромосом.
Все эти причины обусловлены генетически, и поэтому в каждом отдельном скрещивании есть возможность установить главный фактор, определяющий бесплодие.
Атипические митозы в гибридном организме могут начинаться очень рано и приводить к полной депрессии развития. Однако атипичность митозов может быть частичной и не исключает дальнейшего, даже нормального, развития гибридного организма. Однако для полного восстановления плодовитости гибридов требуется полное замещение хромосом одного из исходных видов.
Плодовитость отдаленных гибридов первого поколения часто зависит от выбора линий для скрещивания. Разница в плодовитости объясняется различиями геномов скрещиваемых форм.
Главной причиной, определяющей бесплодие отдаленных гибридов, является нарушение мейоза в гаметогенезе. Нарушение мейоза может быть вызвано рядом генетических факторов.
1. Различие в геномах, что ведет к нарушению равного распределения хромосом в метафазе I.
2. При общем сходстве геномов и равенстве числа хромосом у скрещиваемых видов имеются различия в отдельных аллелях, а также асинаптические гены, препятствующие нормальному спариванию хромосом. Нарушение мейоза может быть вызвано не только генетическими причинами, но и неблагоприятными факторами внешней и внутренней среды. У животных особое значение приобретают гармональные факторы.
При скрещивании видов и цитологическом анализе степени конъюгации хромосом и образования бивалентов и унивалентов в мейозе удается выяснить гомологичность геномов и причины стерильности гибридов, вызванные несовместимостью геномов.
2.9 Способы получения фертильных гибридов
Амфидиплоидия (аллополиплоидия) – способ получения плодовитых отдаленных гибридов, при котором в потомстве сохраняются геномы обоих скрещиваемых видов. Используется для восстановления плодовитости. Как правило, гибриды имеют пониженную плодовитость.
Указанный способ применяют в селекции в том случае, когда преследуют цель получить от одного из скрещиваемых диплоидных видов ценные качества для создания новых сортов, а гибриды первого поколения оказываются стерильными.
Предполагается, что первый отдаленный гибрид, возникший путем удвоения числа хромосом, был обнаружен в 1912 г. Гипотеза получения константных и плодовитых тетраплоидных форм у растений с помощью аллополиплоидии была предложена О. Винге еще в 1917 г.
2.10 Прививки хвойных пород
Прививки растений – это способы получения таких организмов, которые состоят из двух различных индивидуумов. Поэтому в каждой прививке обычно различают две составные части: привой – та часть прививки, которая перенесена на другое растение и срослась с ним, и подвой – часть прививки, на которую перенесен привой и которая его питает. Привой вынужден брать питательные вещества другого растительного организма. Эти новые условия изменяют свойства протоплазмы, а следовательно, и наследственность организма.
2.10.1. Основные способы прививок
Практике известно множество способов прививок, которые сводятся к трем основным:
1) окулировка, или прививка «глазком», – когда привоем служит почка, или «глазок», взятая из средней части побега. Техника прививки этим способом сводится к нанесению на подвое «Т» – образного надреза коры, отчленению «глазка» продольным срезом до коры, помещению «глазка» в подготовленный надрез и наложении повязки;
2) копулировка, или прививка черенком, – способ прививки для размножения наиболее ценных сортов плодовых и форм древесных пород. При нем из однолетних побегов нарезаются черенки с 2–3 почками. На подобранных с одинаковой толщиной черенке привоя и стебле подвоя делают косые срезы так, чтобы их камбиальные слои совпали. При этом для более надежного срастания поверхности среза делают с выступами, углублениями и место сочленения обвязывают. Поскольку у деревьев и кустарников диаметры черенка привоя и стебля подвоя часто не совпадают, прививку делают «в расщеп», «под кору» и другими способами. При прививке в расщеп поперечный срез подвоя расщепляется по диаметру полностью или частично и в него вставляется привой, который косо срезается с двух сторон. При прививке под кору на подвое делают горизонтальный срез под стеблевым узлом, а затем надрезают кору в вертикальном направлении от среза книзу и осторожно отворачивают ее края. На привое срез проводят в виде полуконуса. Затем привои вставляют подкору, зажимают его отворотами коры и обвязывают прививку;
3) сближение, или аблактировка, – способ прививки, при котором привой не отделяется от материнского растения до полного сращивания с подвоем. Способ состоит в том, что в местах подвоя и привоя снимается одинаковая полоска коры с тонким слоем древесины. Затем срезы плотно соединяют и закрепляют повязкой. Для успеха прививки необходимо, чтобы было достигнуто совмещение камбиальных слоев и на стороне привоя, противоположном срезу, имелась почка. Через один-два года, когда компоненты прививки срастутся, привой отделяют от материнского растения.
2.10.2 Способ ментора
И.В. Мичурин показал, что путем прививки и умелого питания растений одной породы пластическими веществами, вырабатываемыми другой породой, можно не только изменить наследственность организма, но и получить в результате настоящие помеси – гибриды.
Метод воспитания под воздействием органического питания широко применялся И.В. Мичуриным и назван им способом ментора.
Способ ментора заключается в прививке в крону молодого (3–5 лет) сеянца черенка с плодоносящего дерева. Сеянец под влиянием привитого на него сорта – ментора в следующие два года приносит плоды. Применяя этот способ, можно не только ускорять, но и повышать плодоношение.
Для наибольшего влияния подвоя на привой и обратно И.В. Мичурин рекомендует регулировать листовую поверхность у привитых компонентов, т.е. удалять часть листьев у того прививочного компонента, который хотят изменить, с тем, чтобы принудить его питаться пластическими веществами другого компонента и таким образом направить развитие того или иного признака в желательную сторону.
Из учения о менторах непосредственно вытекает другой метод, метод вегетативного сближения, который широко использовался и может быть применен для получения гибридов при отдаленной гибридизации. Метод вегетативного сближения основан на том, что если черенки одного вида привить к растению другого (не скрещивающегося половым путем с первым видом), а молодые черенки второго вида привить к первому, привитые компоненты путем взаимовлияния так перестроят свою воспроизводительную систему, что способны будут скрещиваться между собой.
Способы менторов и вегетативного сближения И.В. Мичурин применял главным образом к плодовым растениям, но они вполне могут быть использованы также для других древесных пород и, в частности, для хвойных.
2.10.3 Самопрививки
Самопрививкой у растений называются случаи срастания в естественных условиях (без вмешательства человека) или отдельных частей одного организма, или двух рядом растущих организмов одного и того же вида, а иногда и разных. Самопрививки лиственных древесных пород – явление довольно распространённое. Классический пример весьма легкого срастания ветвей и стеблей одного и того же дерева в местах соприкосновения их можно наблюдать у железного дерева (Раrrоtia persica).
Самопрививки у хвойных деревьев зарегистрированы значительно реже и поэтому они наиболее интересны.
В «Лесном журнале» (1878 г.) приведено описание кедров, выросших на живой сосне. Из ствола сосны, на расстоянии 6 м от земли, с двух сторон выходят в виде сучьев два кедра, имеющие прекрасный рост и густую хвою. Оба кедра ежегодно приносят кедровые шишки. Вначале сосна имела прекрасный рост и в столетнем возрасте достигала 22 м высоты при толщине в диаметре на высоте груди 48 см, а немного выше места сращения – всего 28 см. Диаметр кедров при основании равнялся 16 см. В дальнейшем рост и развитие кедровых сучьев сильно отразилось на росте сосны, которая принимала все более угнетённый вид.
В 1939 г. в газете «Уральский рабочий» (21 февраля) помещена заметка, содержащая в себе описание двух сосново-кедровых гибридов. Один гибрид представляет собой «ствол и сучья до высоты 3–4 метра сосновые, а выше кедровые, с кедровыми шишками и орехами. Второй интересный гибрид – половина ствола от корня – сосна, а выше дерево разветвляется на два: одно ветвление сосна, другое кедр. Обе части имеют разные шишки и семена».
Несколько деревьев – самопрививок сосны с кедром – произрастает сейчас на Урале. За ними наблюдают и они охраняются от повреждений.
Большой научный и практический интерес представило бы скрещивание на описанных прививках сосны с кедром, поскольку описанные самопрививки служат хорошим примером разработанного И.В. Мичуриным метода вегетативного сближения, весьма действенного при отдаленных межвидовых скрещиваниях.
Явление самопрививок наблюдается и у других хвойных пород. Н.П. Кренке описывает случай срастания елей. На двадцатом году жизни росшие рядом стволы срослись в нижних своих частях. Произошло это потому, что камбий в месте повреждения откладывал общие годичные кольца. Таким образом, бывшие раздельными, хотя и тесно прижатые, стволы оказались охваченными общим, наросшим от сращения, кольцом камбия.
У пихты кавказской часто наблюдаются случаи срастания корней. Благодаря такому сращению, у пихты остается после срубки «живой пень». Нормальный пень срубленной пихты, как у большинства хвойных, умирает. В данном же случае пень, получающий органическое питание от соседней пихты через сращенные с ней корни, продолжал откладывать новые годичные кольца, не давая никакой поросли. В свою очередь, пень подкармливал соседку минеральной пищей, т.е. корни пня не только не умерли, но даже давали новые почки. Массовое срастание корней сосен и елей часто можно видеть в чистых еловых и сосновых насаждениях.
Примеры самопрививок, наблюдающиеся среди хвойных древесных пород в надземной и в подземной частях, служат прекрасным доказательством возможности прививок не только внутривидовых, но и межвидовых. Прививки среди хвойных могут быть использованы для размножения редких экземпляров и для получения вегетативных гибридов.
Явления самопрививки у хвойных пород в естественных условиях указывают на возможность срастания этих объектов в условиях искусственных. Способность ветвей и стебля срастаться при соприкосновении, даже без вмешательства человека, давно используется практикой. Существует очень много способов прививки хвойных, которые в деталях несколько отличаются от способов прививок лиственных пород.
2.10.4 Инструменты, употребляемые при прививке
Инструменты, употребляемые при прививках, немногочисленны.
К ним должны быть отнесены: окулировочный нож, нож для копулировки (прививки), садовый нож и садовые ножницы.
Окулировочный нож употребляется главным образом при окулировке. Основные требования, которым должен отвечать окулировочный нож, состоят в том, чтобы верхняя часть лезвия была закруглена и остро отточена. Копулировочный – прививочный нож имеет прямое лезвие и трехгранный, суживающийся книзу черенок.
Садовый нож, служащий для подчистки подвоев, срезки сучков и веток и для сглаживания порезов, делается с закривленным клинком и ручкой. Ручка книзу утолщена и хорошо держится при работе в руке.
Садовые ножницы служат для срезывания черенков с маточного дерева и для подрезки прививаемых частей толщиной больше 2–3 см. Клинок садовых ножниц не должен быть закругленным и остроконечным. Но как бы ни была совершенна конструкция ножниц, они всегда в большей или меньшей степени сдавливают стебель, а потому срезы, в особенности на ветках, предназначенных для прививки, необходимо после ножниц подчищать ножом.
Для предохранения прививок от доступа воздуха, высыхания и загнивания, а также для того, чтобы привой и подвой плотнее прилегли друг к другу, место прививки туго обвязывается мочалом, обязательно влажным. Влажность способствует большей эластичности мочала, которое плотнее прилегает к месту прививки. Чтобы при обвязке прививки не внести какой-либо инфекции, мочало нужно промыть и прокипятить в воде. Поверх мочала накладывается садовый клей. Существует много способов и рецептов приготовления садового вара. Хорошей замазкой является «мастика» из смеси еловой или сосновой смолы, бараньего жира, золы и спирта.
Лучшим варом для замазки прививок хвойных пород считается состав из: канифоли – 1 кг, воска – 0,4 кг, рыбьего жира – 0,25 кг и винного спирта-100 см3 . Этот вар совершенно безвреден для дерева, хорошо намазывается и ложится при употреблении, не стекает от солнечного припека, обладает большой эластичностыо и не трескается от мороза. В некоторых случаях, когда требуется частое ослабление повязки, хорошо применять изоляционную ленту без обмазки ее варом. Для этой же цели прекрасным материалом является медицинский лейкопластырь, который легко достать в любой аптеке.
Комментарии. Во-первых, с помощью прививки гибриды не получают. Получаются химерные организмы, состоящие из 2-х и более частей, каждая из которых обладает собственным генотипом. Гибриды же получаются в результате гибридизации (скрещивания), т.е. в процессе полового размножения а не вегетативного.
Во-вторых, при прививке изменения генотипа, а следовательно и наследственности, не происходит. Может происходить изменение некоторых признаков в пределах нормы реакции, но эти изменения не наследуются.
В-третьих, части химерного растения, полученного в результате прививки, обоюдно обмениваются пластическими веществами, привой синтезирует их в листьях и поставляет подвою, корни подвоя поставляют воду, минеральные соли и некоторые пластические вещества привою. Эти вещества не могут изменить генетической природы компонентов прививки. Даже в описанных выше случаях сосново-кедровых химер каждая из составляющих их частей имела плоды, присущие либо кедру, либо сосне. Смешение признаков не происходило.
2.11 Гибридизация лиственницы
В настоящее время наметились два направления использования гибридизации лиственницы в лесном хозяйстве:
1) отбор среди гибридных растений наиболее быстрорастущих особей, размножение их вегетативным путем с закладкой промышленных плантаций из лучших клонов;
2) получение гибридных семян от наиболее перспективных вариантов скрещивания с использованием их в лесокультурной практике на общих основаниях. Из семян гибридного сорта выращивают сеянцы, саженцы и закладывают культуры, в которых по законам конкуренции и влиянию отбора пойдет процесс самоизреживания.
Первые гибриды от межвидовых скрещиваний в роде Larix Mill, получены еще в XIX в. Примером хороших комбинационных возможностей межвидовых скрещиваний может быть широко известный гибрид между японской и европейской лиственницами (LarixЧeurolepis), впервые обнаруженный в Шотландии в начале XX в., а затем полученный искусственно. В СССР гибриды от этого варианта скрещивания были получены А.С. Яблоковым и А.В. Альбенским, Гибриды отличаются устойчивостью против рака, быстрым ростом, формируют хороший ствол. Работы по контролируемому скрещиванию этих видов проведены в Бельгии, Дании, Швеции и США. Доказано превосходство по росту гибридов первого поколения от скрещивания лиственницы японской с европейской перед родительскими видами в странах Западной Европы.
В СССР этот и другие варианты межвидовых скрещиваний лиственниц используются для получения гибридов в массовом количестве. С этой целью создаются специальные лесосеменные гибридные плантации путем смешанных посадок из двух видов, клоны которых располагаются в рядах так, чтобы обеспечивалось переопыление их между собой. Такие плантации созданы в Литовской ССР, Московской области и др. Лучшие особи отбирают в итоге клопового отбора или испытания в сравнительных посадках, «лиственничных садах» (ларицетумах) корнесобственных растений. Примером такой работы может служить итог первичного изучения и оценки гибридного фонда ВНИИЛМа. Для дальнейшей опытно-производственной проверки и внедрения в промышленные плантации ВНИИЛМ рекомендует следующие гибриды лиственницы.
1. Гибрид Larix deciduaЧLarix leptolepis. Получен А.С. Яблоковым в 1935 г. Из гибридной семьи отобраны три самых лучших дерева, которые в возрасте 35 лет имеют высоту 20 м, диаметр 35 см. Стволы прямые. Гибриды зимостойкие, засухоустойчивые и отличаются очень длительными сроками вегетации хвои по сравнению с родительскими формами.
2. ГибридLarixsibiricaЧLarixdeciduas. Получен А.С. Яблоковым в 1935 г. Из гибридной семьи отобраны три лучших дерева, которые в возрасте 35 лет достигли высоты 18 м, диаметра на высоте груди 38 см. Стволы прямые. Гибриды зимостойкие, засухоустойчивые и отличаются более длительными сроками вегетации хвои по сравнению с родительскими формами.
3. Тройной гибрид LarixleptolepisЧ Larixhibrida№29 (LarixsibiricaЧ Larixdecidua). Получен P. Ф. Кудашевой в 1951 г. Из гибридной семьи отобраны восемь лучших деревьев в качестве маточных, которые в возрасте 28 лет достигли высоты 20 м, диаметра на высоте груди 28 см.
4. Гибрид Larix leptolepis Ч Larix sibirica. Получен P. Ф. Кудашевой в 1951 г. Из гибридной семьи отобраны восемь лучших деревьев в качестве маточных. В возрасте 28 лет высота этих деревьев 20 м, диаметр на высоте груди 28,3 см.
2.12 Морфолого-анатомические маркеры
У хвойных Северного полушария современный этап эволюции связан с гибридизацией и, в частности, интрогрессией, противостоящей процессам специализации. Объединение генетической информации разных видов при «раскачивании» климата в системе стадиал-межстадиал заменяет генерализацию, ставшую невозможной из-за быстрого разнонаправленного изменения условий среды за последние 90–110 тыс. лет. В то же время, формирование рядов эколого-географической изменчивости убыстряется именно благодаря появлению гибридных таксонов. Последние установлены при анализе морфологии генеративных органов, морфолого-анатомических особенностей листа, кариоморфологических характеристик. Особенно широкая вариабельность всех этих признаков почти всегда указывает на гибридность, а отклонения от типичных секционных признаков, например, в положении смоляных каналов хвои (СМК) могут быть настолько существенными, что возникает впечатление межсекционной гибридизации. С другой стороны, гибридизацию внутри видового ряда обнаруживают как по форме и величине генеративных органов, в частности, форме кроющих чешуй у елей секции Casicta, так и по многовершинной кривой вариации степени спирализации хромосом.
У елей аянской группы – дальневосточных Picea ajanensis, P. microsperma, P. komarovii и североамериканской P. sitchensis шишки здесь меняются от коротких цилиндров 4Ч2 см с овальными кроющими чешуями, глянцевых, кофейных (Восточно-Маньчжурские горы и Южный Сихотэ-Алинь) до узких, извитых, блестящих длиной до 12 см на берегу Татарского пролива и узких, более коротких, матово-песочных на юго-востоке о-ва Сахалин и даже до широкояйцевидных 7Ч5 см, кремово-коричневых, блестящих, с прямоугольными кроющими чешуями на Камчатке. Многовершинная кривая степени спирализации хромосом, установленная для елей Камчатки подтверждает гибридизацию здесь P. ajanensis и P. microsperma обусловливающую разброс форм кроющих чешуй. По разбросу форм кроющих чешуй с преобладанием пятиугольной, по некоторой вытянутости лоснящихся шишек и опушению молодых побегов мы вслед за В.Н. Ворошиловым признаем Picea kamtschatkensis самостоятельным гибридогенным таксоном. Однако распространён этот сингамеон и на материке – по Северный Сихотэ-Алинь и – в КНР – бассейн р. Сунгари. На юго-востоке Сахалина у с. Охотское произрастает Picea microsperma без признаков гибридности, которая уже в нескольких десятках километров к северо-западу сменяется сингамеоном P. microsperma Ч P. komarovii. По всему югу Дальнего Востока встречаются популяции плоскохвойных елей, вероятно, мало затронутых гибридизацией и демонстрирующих типовые признаки, установленные В.Н. Васильевым [1]. Для P. komarovii это популяции юго-запада и юга Приморья – Восточно-Маньчжурские горы, Южный Сихотэ-Алинь. Шишки здесь короткие, едва превышающие 4 см, цилиндрические. В то же время, в районе г. Советская Гавань у гибрида елей мелкосеменной и Комарова длина шишек в среднем 63,58±1,00 мм, форма кроющих чешуй преимущественно пятиугольная. В 300 км к северу (район пос. Де-Кастри) шишки ели камчатской достигают иногда и 130 мм в длину при средней длине около 80 мм. Но и здесь встречаются популяции ели с короткими шишками, природа которых пока не ясна.
В ареале Pinus densiflora в Хасанском и Шкотовском районах Приморского края обнаружены изолированные популяции с СМК, расположенными преимущественно в паренхиме числом до 10. У елей с верховьев р. Комиссаровка (Пограничный район Приморья) до 9 СМК лежит в паренхиме листа, хотя для Pinus Ч funebris типично 1–2 (3) канала в паренхиме при абсолютном преобладании периферических каналов; не менее 90% хвоинок имеют периферические СМК. Таким образом, доказано наличие в Приморье и сопредельных странах более молодых, чем возникшая к середине миоцена P. Чx funebris, гибридных сосен P. Чx densi-thunbergii (ранее известна из Японии) и P. Ч funebri-thunbergiana. Их разрозненные, изолированные популяции маркируют рефугиумы наиболее теплолюбивых неморальных форм, устойчивых к периодическим засухам которые находятся в урочищах с наиболее теплым в течение всего плейстоцена микроклиматом, мелкоформенным рельефом, глубокими речными долинами, что обусловливает наличие мозаики экотопов, часть которых благоприятна для P. Чx densi-thunbergii или P. x funebri-thunbergiana в любую фазу ротации климата системы стадиал-межстадиал.
Морфолого-анатомические маркёры гибридизации 1) показали существенно большее разнообразие на Дальнем Востоке родственных видов в предшествующие геологические эпохи как на рубеже стадиалов, так и в последнем стадиале, 2) могут помочь определить местоположение рефугиумов, 3) вскрыть масштабы смещения и перекрывания ландшафтных зон, 4) позволяют обнаружить урочища с оптимальным в стадиалы и наиболее тёплым сейчас микроклиматом, 5) отражают массовую замену генерализационных процессов гибридогенной эволюцией именно в стадиалы, что может быть показателем некоторой регрессивности эволюционного процесса на современном этапе.
Заключение
Меж- и внутривидовая гибридизация – один из наиболее перспективных методов в программах селекционного улучшения древесных растений. При этом возможен, в частности, гетерозисный эффект по самым разнообразным признакам: скорости роста, мощности, урожайности. Другой селекционный выход заключается в получении новых комбинаций признаков, отборе в гибридном потомстве особей с высоким качеством древесины, устойчивых к болезням, повреждениям морозами. Гибридизация совместимых видов часто обеспечивает возможность массового получения быстрорастущих или высокопродуктивных гибридов первого поколения, которые непосредственно могут быть использованы в лесном хозяйстве. Искусственная гибридизация (в том числе на основе отдаленных скрещиваний) в сочетании с отбором в последующих поколениях является способом создания новых гибридных сортов деревьев.
В рамках популяционной селекции можно говорить о межпопуляционной (и межрасовой) гибридизации с целью повышения генетического разнообразия получаемого потомства путем комбинирования генетического материала (генофонда) из различных популяций, например, в специально создаваемых гибридно-семенных плантациях. При этом, как уже отмечалось (см. выше), обеспечивается своего рода искусственное восстановление «потока генов» между ныне пространственно изолированными, а ранее, возможно, обменивавшимися своим генофондом популяциями. Это в особой степени относится к регионам, в пределах которых популяции характеризуются пониженной фенотипической и генетической изменчивостью.
Литература
1. Тренин В.В. Введение в цитоэмбриологию хвойных. Петрозаводск, 1988.
2. Ирошников А.И. Селекция хвойных пород Сибири. Институт леса и древесины. Красноярск, 1978.
3. Ирошников А.И. Изучение полиморфизма популяций древесных пород. Лесная генетика, селекция и семеноводство. Петрозаводск: Карелия, 1970.
4. Любавская А.Я. Лесная селекция и генетика. М.: Лесн. Пром-сть, 1982.
5. Политов Д.В. Некоторые итоги популяционно-генетического изучения хвойных Сибири. Институт общей генетики им. Н.И. Вавилова РАН, Москва, Россия, 1999.
6. В.П. Путенихин, Г.Г. Фарукшина. Методы сохранения генетической гетерогенности при создании искусственных популяций лесообразующих видов.Ботанический сад-институт УНЦ РАН, Республика Башкортостан, Уфа, 2002.