Оптимизация воздействия на микробиоценоз дерново подзолистой почвы комплекса органо-минеральн
СОДЕРЖАНИЕ: Курсовая работа Оптимизация воздействия на микробиоценоз дерново – подзолистой почвы комплекса органо-минеральных удобрений при выращивании турнепса.Курсовая работа
Оптимизация воздействия на микробиоценоз дерново – подзолистой почвы комплекса органо-минеральных удобрений при выращивании турнепса.
Владимир 2010г.
Введение
Биологический круговорот элементов включает поглощение растениями из атмосферы углекислоты и кислорода, а из почвы – азота,кислорода, калия, кальция, магния и многих других элементов; построение из них растительных организмов; разложение отмерших растительных остатков, освобождение элементов; вовлечение этих элементов в новый биологический круговорот.
Образование почвы возможно только под воздействием живых организмов, поселяющихся на материнских породах. В почвообразовании участвуют три группы живых организмов: низшие - организмы (микроорганизмы, лишайники), зеленые растения и животные. Их совместная деятельность превращает породу в новое природное тело- почву.
Микроорганизмы являются первыми поселенцами на любой горной породе. Их несколько миллиардов на 1 г почвы. Их количество и состав варьирует в зависимости от природных условий, характера других факторов почвообразования. В северных районах их меньше и они распространены на небольшую глубину. На юге – их больше и захватывают большую толщу. Окультуренные почвы содержат больше микроорганизмов.
Микроорганизмы разлагают органические остатки, превращая их в гумус. Превращают сложные органические соединения в минеральные соли, доступные для растительности, усваивают атмосферный азот и снабжают им высшие растения, синтезируют сложные органические соединения, строя из них свое тело. Участвуют во всех окислительно - восстановительных процессах в почве, изменяя степень окисленности различных органических и минеральных соединений. Таким образом, почти все звенья почвообразовательного процесса связаны с жизнедеятельностью микроорганизмов. Все эти процессы микроорганизмы осуществляют свою деятельность при помощи ферментов.
Бактерии – наиболее распространенная группа микроорганизмов. Минимальное их количество содержат подзолы, максимальное – черноземы. Распространены бактерии в верхнем слое почв неравномерно. Больше их около корней высших растений и в 2-3 мм от них - это прикорневой слой почвы - ризосфера. Количество бактерий здесь в 20 раз больше среднего числа. Около корней каждого растения развивается специфическая микрофлора. Разные по форме: палочки, кокки, извитые по форме. Разные по способу питания: гетеротрофные и автотрофные. Аэробные и анаэробные.
К группе гетеротрофов относится большинство почвенных бактерий. Очень разнообразны по физиологическим функциям: одни разлагают белки, другие жиры, углеводы.
Гетеротрофные анаэробные бактерии вызывают процессы брожения углеводов. Некоторые представители рода Clostridium обуславливают масляно-кислое брожение углеводов.
Аммонификация – это процесс распада белков вначале до аминокислот, а далее аминопептидазы осуществляют следующие реакции:
гидролитическое дезаминирование –образуется оксикислоты и аммиак
Процесс аммонификации имеет огромное значение для почвообразования, так как ему почвы обязаны накоплению минерального и азота, доступного растениям. Накопление NН3 в почве наблюдается в том случае, когда отношение углерода и азота в разлагающимся материале превышает 20. Если оно меньше, то азот идет на построение биомассы микроорганизмов. Поэтому наличие в почве большого количества неразложившегося органического вещества ведет к торможению накопления азота в почве.
Азотофиксирующие бактерии наиболее интенсивно проявляют свою деятельность при оптимальной температуре. Имеются две группы бактерий: свободноживущие аэробы и анаэробы и симбиотические клубеньковые бактерии. Бобовые растения выделяют безазотистые органические вещества (сахарин), а бактерии окисляют эти вещества и сопряженно фиксируют молекулярный азот воздуха. Нитрифицирующие бактерии вызывают биохимические процессы окисления восстановленных форм азота до азотной кислоты. К нитрификаторам относят представителей родов Nitrosomonas, Nitrosococcus, Nitrosolobus, Nitrosospira, Nitrosovibrio, Nitrobacter. Процесс нитрификации идет в две фазы, за каждую из которых ответственны свои возбудители. В Первую фазу осуществляется окисление солей аммония до солей азотистой кислоты (нитритов), во вторую фазу нитриты окисляются в нитраты
NН4 + +1/2 О2 = NО2- + Н2О + 2 Н+
NO2- 2 - + 1/2 O2 = NO2- 3
По типу питания нитрификаторы являются хемолитоавтотрофы, то есть используют энергию, которую получают при окислении аммиака до нитрита для ассимиляции углекислого газа.
Большую роль относят к бактериям, окисляющим соединения серы, например сероводород, тиосульфат, молекулярную серу, бактериям, осуществляющим окисление железа и марганца, которые также используют энергию окисления в форме АТФ для автотрофной ассимиляции углекислого газа.
Актиномицеты или «лучистые грибки», являются переходной формой между бактериями и грибами. Это аэробные,гетеротрофные организмы. Разлагают органику. Играют большую роль в почвообразовании. Хорошо развиваются в нейтральной и слабощелочной среде. Часто выделяют летучие вещества . Запах земли обусловлен их присутствием.
Грибы . Нельзя не отметить большую роль грибов, которые лучше развиваются в закисленных почвах. Все они гетеротрофы. Обладают широким комплексом гидролитических ферментов, посредством которых существляют разложение всех видов органических веществ. Разлагают устойчивые к гидролизу и окислению такие соединения как лигнин, фенолы, хиноны, ароматические углеводороды.
Водоросли . В почве распространены диатомовые, сине-зеленые и зеленые водоросли. Количество их зависит от увлажнения почвы. Автотрофы. Синтезируют органику путем фотосинтеза, а попадая в глубокие слои горных пород, могут использовать для питания органические вещества. Водоросли обогащают почву органическим веществом, легко разлагающимся микроорганизмами. Участвуют в процессах выветривания горных пород.
Лишайники . Включают в себя автотрофный фикобионт и гетеротрофный микобионт. Разрушают горные породы. Выделяют особые кислоты, разрушающие минералы горных пород. Из продуктов разрушения используют многие элементы для своего питания. Слоевище лишайников прочно срастаются с поверхностью пород. Слоевище при отмирании и высыхании отрывает от поверхности тонкую ее пленку – под лишайником образуется тонкий слой рыхлой, выветренной горной породы.
Зеленые растения . Общая масса организмов на планете составляет 17-18 тонн. Масса растений в 10 – 100 тыс. раз превышает массу животных. Ведущее место в синтезе почвенной органики принадлежит растениям.
Различные группы растений обусловливают неодинаковый ход биологического круговорота. Низшие растения имеют небольшую продолжительность жизни и, следовательно, определяют быстрое обращение элементов в биологическом круговороте. Высшие растения имеют развитую корневую систему, обеспечивающую большую площадь соприкосновения организма с почвой. Круговорот осуществляется в течение одного года – у травянистой растительности и в течение нескольких лет, (десятков, сотен, тысяч) – у древесной. При этом части элементов удерживаются растительным организмом. В природе часто наблюдается сочетание рассматриваемых групп растений.
Количество органического вещества, ежегодно синтезируемого растениями, его качественный состав, интенсивность биологического круговорота обусловлены типом растительной формации: арктическа тундра-4-5 ц/га; хвойные леса северной тайги - 50 ц/га; южной – 4-5 ц/га; широколиственные леса – 200 ц/га; ковыльные степи – 70 ц/га; луговые степи – 100 ц/га; опустыненные степи – 50 ц/га; пустыни – 5 ц/га.
Растительные организмы содержат от 20 до 90 % воды. В сухой массе углерода содержится 45%, кислорода – 42%, водорода – 6,5%, азота– 1,5%, золы – 5%. В составе золы входят почти все элементы периодической системы элементов. Но чаще всего это – калий, кальций, сера, фосфор, кремний, магний. В древесине много калия и магния ,в травянистой растительности много кремния. Растительные остатки содержат: растворимый сахар, крахмал, гемицеллюлозу, клетчатку, лигнин, белковые вещества, жиры и другие соединения. Больше всего клетчатки - 20-40%, а в древесине – до 60%. Здесь же больше чем в траве лигнина – (20-30%). Белка в растениях содержится от 0,6 до 1,0% в древесине и до 15% - в травянистых видах. Крахмала много в зернах и клубнях (70% и 20% соответственно).
Микроскопических животных в почве больше всего, более 2 млрд. на 1 га. площади. Велика роль в почвообразовании червей, а также млекопитающих, живущих в почве, прокладывающих в почве ходы диаметром от 4 до 12 см., перемешивающие почву на разные глубины, в основном на глубину до 1 метра, выделяющие ферменты, органические кислоты, увеличивающие собой биомассу почвы.
Животные и их роль в почвообразовании. Микроскопических животных в почве больше всего, более 2 млрд. на 1 га. площади. Велика роль в почвообразовании червей, а также млекопитающих, живущих в почве, прокладывающих в почве ходы диаметром от 4 до 12 см., перемешивающие почву на разные глубины, в основном на глубину до 1 метра, выделяющие ферменты, органические кислоты, увеличивающие собой биомассу почвы. Живые организмы — обязательный компонент почвы. Количество их в хорошо окультуренной почве может достигать нескольких миллиардов в 1 г почвы, а общая масса — до 10 т/га. Основная их часть — микроорганизмы. Доминирующее значение принадлежит растительным микроорганизмам (бактерии, грибы, водоросли, актиномицеты).
Богатая микроорганизмами почва склеивается минеральными и органическими коллоидными частицами в мелкие комочки, которые неплотно прилегают друг к другу, что позволяет воздуху проникать вглубь почвы, а воде не задерживаться на поверхности и смачивать почву. Богатая гумусом глина рассыпается на мелкие комочки.
Структурность почвы - важнейшее условие синтеза гумуса, наращивания плодородия почвы, ее здоровья.
Ходы микроскопических и дождевых червей, полости отмерших корней растений также улучшают аэрацию и проницаемость почвы. Внесение извести в тяжелую глинистую кислую почву тоже улучшает ее проницаемость и структуру.
Микроорганизмы почвы весьма разнообразны по составу и биологической деятельности. Здесь распространены бактерии, актиномицеты, грибы,водоросли, простейшие. Суммарная масса микроорганизмов только в
поверхностном горизонте достигает нескольких тонн на гектар. Численность
микроорганизмов измеряется миллиардами в 1 г почвы. В целом для планеты
масса почвенных микроорганизмов определяется в 10^8-9 т, т. е. составляет
0,01-0,1% от всей биомассы суши.
Бактерии - это одноклеточные организмы размером в несколько микрометров.
По характеру поглощения углерода выделяют автотрофные бактерии,
усваивающие углерод из воздуха, и гетеротрофные, получающие углерод из
готовых органических соединений. По отношению к азоту лишь часть бактерий
автотрофна, т. е. способна усваивать этот элемент из воздуха.
Автотрофные бактерии поглощают углерод из углекислоты; этот процесс
эндотермический, требующий затраты дополнительной внешней энергии. В
качестве таковой бактерии используют энергию окисления некоторых
минеральных соединений. Этот процесс получил название хемосинтеза.
Примером осуществления хемосинтеза является деятельность нитрифицирующих
бактерий. Под нитрификацией понимают процесс биохимического окисления
аммиака до азотной кислоты. О количественном масштабе процесса
нитрификации можно судить по тому, что за один год деятельности
нитрифицирующих бактерий может образоваться до 300 кг солей азотной
кислоты на 1 г почвы.
Аналогично происходит хемосинтез у других нитрифицирующих бактерий.
Источником энергии для поглощения углерода из углекислого газа могут
служить реакции окисления сероводорода, тиосоединений серы, соединений Fe
(II), Mn (II) и т. д. Накопление сульфатов в результате деятельности
серобактерий в приповерхностном слое почвы достигает 200-250 кг на 1 г
почвы.
Определенные группы бактерий обладают способностью поглощать молекулярный
азот из воздуха. Этот процесс получил название фиксации азота. Нехватка
азота в почве сдерживает развитие растительности, ограничивает возможности
сельскохозяйственного использования почвы. Значение азотофиксирующих
бактерий чрезвычайно велико, так как только благодаря их деятельности для
всей остальной массы живых организмов становится доступным атмосферный
азот.
Гетеротрофные бактерии поглощают необходимый углерод из готовых
органических соединений, разлагая сложные соединения на простые. Благодаря
их деятельности осуществляется грандиозный процесс разрушения
колоссального количества мертвого органического вещества, ежегодно
поступающего в почву, и освобождение химических элементов, прочно
связанных в составе органических остатков.
Актиномицеты - лучистые грибы. Их используют в качестве источника углерода
разнообразные органические соединения. Они могут разлагать клетчатку,
лигнин, перегнойные вещества почвы. Учавствуют в образовании гумуса.
Актиномицеты лучше развиваются в почвах с нейтральной и слабощелочной
реакцией, богатых органическим веществом и хорошо обрабатываемых. К
актиномицетам относят близкие к ним проактиномицеты, микобактерии,
микромоноспоры и микоккоки.
Среди почвенных микроорганизмов исключительно важное значение принадлежит
грибам. Большая часть грибов состоит из ветвящихся нитей (гиф), образующих
тело гриба (мицелий). Наиболее распространены плесневые грибы, а в лесных
почвах гриб - мукор. Грибы разрушают клетчатку и лигнин, участвуют в
разложении белков. При этом образуются органические кислоты, увеличивающие
почвенную кислотность и влияющие на преобразование минералов. Так же как
актиномицеты, грибы преимущественно являются аэробами.
Мицелий грибов часто развивается на корнях растений и даже в клетках
высших зеленых растений. Подобный симбиоз высших растений с грибами
называется микоризой. В этом симбиозе мицелий гриба выполняет функции
всасывающего аппарата корневой системы, обеспечивая растения водой и
пищей. В силу того, что грибы усваивают питательные вещества
непосредственно из органических соединений, микориза обеспечивает развитие
растений на почвах, богатых слаборазложившимися растительными остатками. В
свою очередь, мицелий грибов использует для питания углеводы и некоторые
органические кислоты, поступающие из листьев в корни растений.
Водоросли распространены во всех почвах, главным образом в поверхностном
слое. Содержат в своих клетках хлорофилл.
В болотных почвах и на рисовых полях водоросли улучшают аэрацию, усваивая
растворенный СО[2] и выделяя в воду кислород.
Водоросли активно участвуют в процессах выветривания пород и в первичном
процессе почвообразования.
Лишайники не относятся к микроорганизмам, но поскольку они представляют
собой сложное симбиотическое образование гриба и водоросли, целесообразно
рассмотреть их участие в почвообразовании. Лишайники поселяются как на
органическом веществе, так и на горных породах. Особый интерес
представляет их деятельность на горных породах. Воду и углерод лишайники
получают из атмосферы, а другие химические элементы - за счет разрушения
минералов.
Помимо растительных организмов в почве распространены простейшие животные
организмы. Это преимущественно корненожки, жгутиковые и реснитчатые
инфузории, но роль их в почвообразовании недостаточно выяснена.
Животные организмы представлены простейшими (жгутиковые, корненожки, инфузории), а также червями. Довольно широко распространены в почве моллюски и членистоногие (паукообразные, насекомые).
Почвенные организмы разрушают отмершие остатки растений и животных, поступающие в почву. Одна часть органического вещества минерализуется полностью, а продукты минерализации усваиваются растениями, другая же переходит в форму гумусовых веществ и живых тел почвенных организмов.
Некоторые микроорганизмы (клубеньковые и свободноживущие азотфиксирующие бактерии) усваивают азот атмосферы и обогащают им почву.
Почвенные организмы (особенно фауна) способствуют перемещению веществ по профилю почвы, тщательному перемешиванию органической и минеральной части почвы.
Важнейшая функция почвенных организмов — создание прочной комковатой структуры почвы пахотного слоя. Последнее в решающей степени определяет водно-воздушный режим почвы, создает условия высокого плодородия почвы.
Наконец, почвенные организмы выделяют в процессе жизнедеятельности различные физиологически активные соединения, способствуют переводу одних элементов в подвижную форму и, наоборот, закреплению других в недоступную для растений форму.
В обрабатываемой почве функции почвенных организмов сводятся к поддержанию оптимального питательного режима (частичное закрепление минеральных удобрений с последующим освобождением по мере роста и развития растений), оструктуриванию почвы, устранению неблагоприятных экологических условий в почве.
В интенсивном земледелии экологические условия могут иногда в решающей степени определять эффективное плодородие почвы. В ней существуют тесные многообразные связи между всеми почвенными организмами. Причем вся эта система находится в состоянии непрерывно изменяющегося равновесия. Одни группы микроорганизмов предъявляют простые требования к пище, другие — сложные. Между одними группами существуют симбиотические (взаимно полезные) связи, между другими — антибиотические. Микроорганизмы в последнем случае выделяют в почву вещества, подавляющие развитие других микроорганизмов.
Практическое значение имеет способность некоторых микроорганизмов оказывать губительное действие на представителей фитопатогенной микрофлоры.
Усилить активность желательных микроорганизмов можно путем внесения в почву органического вещества. В этом случае отмечается вспышка в развитии почвенных сапрофитов, которые, в свою очередь, стимулируют развитие микроорганизмов, угнетающих фитопатогенные виды. Для нормального функционирования почвенных организмов необходимы прежде всего энергия и питательные вещества. Для подавляющего большинства микроорганизмов такой источник энергии — органическое вещество почвы. Поэтому активность почвенной микрофлоры главным образом зависит от поступления или наличия в почве органического вещества.
Для оценки деятельности почвенной биоты используют показатель «биологическая активность почвы». Под биологической активностью понимают, в одних случаях общую биогенность почвы, определяемую, как правило, подсчетом общего количества почвенных микроорганизмов. Если иметь в виду несовершенство методик, применяемых в этом случае, и малую кратность определений во времени, то результаты анализа дают примерную картину биологической активности почвы.
Другая точка зрения относительно методов определения биологической активности почвы заключается в учете результатов деятельности почвенных организмов. Особенно важен такой подход в агрономии. Однако привести к общему знаменателю исключительно многообразную деятельность почвенной флоры и фауны методически непросто.
Наиболее универсальный показатель деятельности почвенных организмов — продуцирование ими углекислого газа. Поэтому учет выделяемого почвой углекислого газа — первостепенный из других биохимических способов определения биологической активности почвы.
Биологический круговорот в почве осуществляется с участием разных групп микроорганизмов. В зависимости от типа почвы содержание икроорганизмов колеблется. В садовых, огородных, пахотных почвах их насчитывается от одного миллиона до нескольких миллиардов микроорганизмов в 1 г почвы. В почве каждого садового участка присутствуют свои микроорганизмы.
Они участвуют своей биомассой в накоплении органического вещества почвы. Они выполняют огромную роль в образовании доступных форм минерального питания растений.
Исключительно велико значение микроорганизмов в накоплении биологически активных веществ в почве, таких как ауксины, гиббереллины, витамины, аминокислоты, стимулирующие рост и развитие растений.
Микроорганизмы, образуют слизи полисахаридной природы, а также большое количество нитей грибов, принимают активное участие в формировании структуры почвы, склеивании пылеватых почвенных частиц в агрегаты, чем улучшают водно-воздушный режим почвы.
Биологическая активность почвы, численность и активность почвенных микроорганизмов тесно связаны с содержанием и составом органического вещества. В тоже время с деятельностью микроорганизмов тесно связаны такие важнейшие процессы формирования плодородия почв, как минерализация растительных остатков, гумификация, динамика элементов минерального питания, реакция почвенного раствора, превращения различных загрязняющих веществ в почве, степень накопления ядохимикатов в растениях, накопление токсических веществ в почве и явление почвоутомления. Велика санитарно-гигиеническая роль микроорганизмов и в трансформации и обезвреживании соединений тяжелых металлов.
Перспективным направлением восстановления и поддержания плодородия и биологической интенсификации земледелия считается применение продуктов переработки органических отходов с участием-вермикомпостов дождевых червей, находящихся в симбиозе с микроорганизмами.
В естественных почвах разложение опада осуществляют дождевые черви, копрофаги и другие организмы. Но в этом процессе участвуют и микроорганизмы. В кишечнике червей для них создаются более благоприятные условия для выполнения любых функций, чем в почве. Дождевые черви в союзе с микроорганизмами превращают различные органические отходы в высокоэффективные биологические удобрения с хорошей структурой, обогащенные макро- и микроэлементами, ферментами, активной микрофлорой, обеспечивающей пролонгированное (длительное, постепенное) действие на растения.
Обеспечивая развитие микроорганизмов в почве, вы, повышаете урожай и улучшаете его качество. Ведь микроорганизмы развиваются, т.е. делятся каждые 20-30 мин и при наличии достаточного питания образуют большую биомассу. Если бык весом 500кг за сутки образует 0,5 кг биомассы, а 500 1 кг, то 500 кг микроорганизмов за сутки создают 5т-кг растений биомассы. На площади 1 га в результате жизнедеятельности почвенных микробов в течение года выделяется 7500м3
углекислоты. А углекислота необходима и как источник углеродного питания растений и для растворения труднодоступных солей фосфорной кислоты и превращения фосфора в форму доступную для питания растений. Т.е. там, где хорошо работают микроорганизмы, нет необходимости во внесении фосфорных удобрений. Но сами микроорганизмы нуждаются в органическом веществе. В балансе органического вещества почвы велика роль культурных растений. Накоплению гумуса в почвах способствует многолетние травы, особенно бобовые. После их уборки в почве остается фитомасса, которая обогащена азотом за счет фиксации его клубеньковыми бактериями из воздуха. Пропашные и овощные культуры (картофель, капуста и др.) уменьшают содержание гумуса в почве, т.к. оставляют в почве небольшое количество растительных остатков, а применяемая система глубокой обработки почвы обеспечивает интенсивное поступление в пахотный слой кислорода и, как следствие, обеспечивает сильную минерализацию органического вещества, т.е. его потерю.
Характеристика почвы
Дерново-подзолистые почвы формируются в результате двух противоположено направленных процессов почвообразования, таких как подзолистый и дерновый. Этот тип почв формируется под хвойношироколиственными, мохотравянистыми и травянистыми лесами в условиях промывного водного режима.
Физические свойства.
В связи с более высоким содержанием гумуса дерново-подзолистые почвы имеют более низкую плотность верхнего слоя, они лучше оструктурены по сравнению в подзолистыми, у них выше пористость. В целом естественное плодородие этих почв выше, чем у подзолистых и глееподзолистых. Они являются преобладающими в пахотном фонде почв таежно-лесной зоны.
Для некоторых почв подзолистого типа большое значение имеет их тепловой режим.
В восточных районах, в условиях континентального климата и недостатка тепла, темные и богатые гумусом дерново-подзолистые и дерново-лесные почвы плохо нагреваются и не обеспечивают корневую систему растений необходимым количеством тепла, что приводит к недозреванию некоторых посевных культур.
Таким образом, наиболее характерными свойствами, присущими почвам дерново-подзолистого типа, являются следующие.
1. Почвы дерново-подзолистого тина не насыщены основаниями, т. е. они содержат в себе то или иное количество обменных ионов водорода и алюминия.
2. В связи с ненасыщенностью основаниями эти почвы обладают кислыми свойствами.
3. В результате процессов минерализации органического вещества и выщелачивания эти почвы обеднены гумусом и минеральными легко подвижными соединениями.
4. Почвы дерново-подзолистого типа заключают в себе сравнительно небольшие количества необходимых для растений питательных веществ.
5. Эти почвы в большинстве случаев обладают слабо выраженной, непрочной структурой, вследствие чего они способны легко распыляться, заплывать при увлажнении, образовывать корку при высыхании и уплотняться.
6. Почвы дерново-подзолистого типа отличаются резко выраженной дифференциацией
7. Почвы дерново-подзолистого типа отличаются резко выраженной дифференциацией почвенного профиля на составляющие их генетические горизонты, причем гумусовый горизонт в большинстве случаен имеет малую мощность.
8. В составе гумуса видное или первое место принадлежит фульвокислотам, относительное количество которых тем больше, чем резче выражен подзолистый процесс. При нарастании дернового процесса увеличивается количество гуминовых веществ.
9. В подзолистых почвах ведущее место принадлежит грибам.
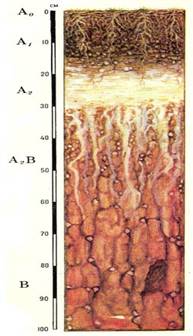
Строение почвенного профиля
Особенность строения почвенного профиля дерново-подзолистых почв состоит в том, что выделяется четкая дифференциация почвенной толщи, которая составляет ее генетические горизонты. Основных ярко выраженных горизонтов четыре: дерновый, или перегнойно-аккумулятивный (A1), подзолистый (А2) иллювиальный горизонт (B) и материнская, или почвообразующая, порода (С). Таю же на поверхности данной почвы всегда находится лесная подстилка или войлок (Aо).
Лесная подстилка Ао
Она состоит из растительных остатков различной степени разложении, очень часто переплетенных гифами грибов. Мощность ее обычно колеблется от нескольких миллиметров до нескольких сантиметров.
Дерновый, или перегнойно-аккумулятивный, горизонт А1
Этот горизонт окрашен сверху в темно-серый цвет находящимся здесь перегноем. С углублением по мере уменьшения гумуса окраска его становится несколько светлее. В верхних частях горизонт состоит из рыхло залегающих комков, все промежутки между которыми заполнены живыми и мертвыми корнями и корневищами, скрепляющими комки о дернину.
Подзолистый, или элювиальный горизонт (А2)
Горизонт является тем почвенным слоем, в котором наиболее резко выражено влияние подзолообразовательного процесса. Он сильно выщелочен, обеднен перегноем и содержит в себе повышенное количество кремнезема в виде тончайшей белой кремнеземной пыли. Эта пыль вместе со светлым гумусом придает горизонту белую или светло-серую окраску.
В зависимости от степени оподзоленности этот горизонт имеет различную мощность (от нескольких до десятков сантиметров). Чаще всего он бесструктурный, лишь в отдельных случаях приобретает отчетливо выраженную пластинчатую или листоватую структуру, которая в производственном отношении, однако, не представляет собой никакой ценности.
Будучи сильно выщелоченным и обедненным органо-минеральными коллоидами, элювиальный горизонт является самым бесплодным слоем дерново-подзолистой почвы.
Иллювиальный горизонт В
Иллювиальный горизонт частично закрепляет выносимые в процессе подзолообразования из верхних горизонтов вещества. Здесь коагулируют и задерживаются коллоидальные гидраты описи железа, алюминия, частично двуокиси кремния, перегнойные вещества, а так же глинистые суспензии. Вследствие обогащенности железом и перегноем иллювиальный горизонт обычно имеет красно-бурую окраску и, будучи пропитан и сцементирован коллоидальными частичками, отличается повышенной плотностью.
С глубиной количество новообразований заметно убывает, почва приобретает более однородную окраску и постепенно переходит в материнскую породу, еще не измененную явным образом почвообразовательными процессами.Может подразделяться над подгоризонты В1 В2, В3.
Материнская, или почвообразующая, порода С — самый нижний горизонт. Эта порода послужила исходным материалом для образования данной почвы. Как правило, горизонт С слабо затронут почвообразовательным процессом, поэтому он всегда легко отделяется от других вышележащих почвенных горизонтов.
Профиль дерново-подзолистой почвы очень изменчив по мощности и степени выраженности отдельных генетических горизонтов.
Очень часто переход между горизонтами бывает довольно резням, в некоторых же случаях настолько незаметен, что трудно бывает даже провести границу между ними.
Состав и свойства
Химический состав.
Состав и свойства дерново-подзолистых почв во многом зависит от дернового почвообразовательного процесса, в результате которого накапливается гумус, основания и элементы питания для растений.
Содержание гумуса в гумусовом горизонте суглинистых разновидностей -3-6%, в песчаных и супесчаных - 1.5-3%. Сравнительная бедность дерново-подзолистых почв перегноем объясняется главным образом тем, что при благоприятных условиях увлажнения распад растительных остатков происходит энергично, не прекращаясь даже в летние, наиболее сухие месяцы. Органическое вещество сосредоточено в основном в самом поверхностном слое, а поскольку этот слой отличается малой мощностью, не превышающей часто S—12 см, то и абсолютное содержание перегноя в описываемых почвах весьма ограничено. Содержание гумуса резко снижается с глубиной в горизонте А2 составляет 0.3-0.5%. Важно отметить, что в составе гумуса преобладаю фульвокислоты над гуминовыми.
В тех почвах, в которых подзолистый процесс ослаблен, а дерновый преобладает, мощность гумусного горизонта возрастает, увеличивается количество гумуса в поверхностном горизонте и в целом по профилю почвы.
Развитие подзолистого процесса под лесной растительностью проходит с различной интенсивностью. В некоторых случаях, например па карбонатных породах, под лиственными насаждениями, этот процесс протекает настолько слабо, что в почвенном профиле или не происходит никаких перемещений минеральной части, или эти перемещения чрезвычайно малы. Подзолообразовательныи процесс влияет на химический состав почвы и вносит изменения в ее механический состав.
верхние горизонты дерново-подзолистых почв по сравнению с материнской породой (горизонт С) обеднены мельчайшими частичками (0,001 мм). В то же время содержание крупных фракций в верхних горизонтах выше, чем в материнской породе. Иллювиальный горизонт (В) более обогащен по сравнению с материнской породой и с вышележащими горизонтами самыми мелкими фракциями (0,001 мм).
Понятно, что такого рода перераспределение вещества по почвенной толще неизбежно сказывается и на физических свойствах почвы: верхние горизонты несколько опесчаниваются, утрачивают связность, делаются более рыхлыми и бесструктурными; нижние же слои, наоборот, уплотняются, становятся связными и трудно-проницаемыми как для воздуха, так и для воды.
Однако такая закономерность обнаруживается только в почвах суглинистых и глинистых, т. о. в почвах тяжелого механического состава и с ярко выраженным подзолистым процессом
В супесчаных и отчасти в песчаных почвах нередко наблюдается несколько иная картина: в верхнем горизонте происходит некоторое увеличение количества мелких частиц и в то же время соответственное относительное уменьшение наиболее крупных фракций.
В составе поглощенных катионов дерново-подзолистых почв выделяются среди поглощенных катионов в поглощающем почвенном комплексе поглощенный водород и алюминий. Наибольшая ненасыщенность наблюдается в самых верхних, оподзоленных горизонтах; здесь поглощенный Н нередко может преобладать над поглощенными Са и Мд.
По мере углубления но почвенному профилю соотношение поглощенных катионов заметно изменяется: количество водородных попов и алюминия прогрессивно убывает, а Са и Мд возрастает и уже в горизонте С, не затронутом еще почвообразовательными процессами, поглощенный Н сходит почти на нет, и в поглощающем комплексе оказываются одни лишь металлические катионы.
Таким образом, в результате оподзоливания верхние горизонты обедняются поглощенными основаниями, обогащаясь в то же время водородными ионами и обменным алюминием. Наличие обменных катионов водорода придает почве кислотные свойства.
Весьма характерно, что степень кислотности каждого горизонта находится в полном соответствии с его насыщенностью основаниями; в самых верхних горизонтах, сильно вы щелоченных и насыщенных водородными ионами, обнаруживается повышенная кислотность. При переходе к нижним горизонтам, менее выщелоченным, кислотность постепенно падает, и в материнской породе она сменяется реакцией, близкой к нейтральной, или в случае карбонатности породы — слабощелочной.
Физико-химические свойства.
Емкость катионного обмена в дерново-подзолистых почвах выше, чем в подзолистых, в связи с более высоким содержанием гумуса. В гумусовом слое суглинистых и глинистых почв она достигает 15-20 мг-эке на 100 г, в песчаных и супесчаных — только 4-10 мг-эке. В составе поглощенных катионов — Са2+ , Mg2+, H+ и А13+. Степень насыщенности основаниями — 40-70%. Обменные катионы водорода и алюминия обусловливают кислую реакцию среды (рНкс! обычно варьирует в пределах 3,3-5,5)7.
Физический свойства.
В связи с более высоким содержанием гумуса дерново-подзолистые почвы имеют более низк/ю плотность верхнего слоя, они лучше острукгурены по сравнению в подзолистыми, у них выше пористость. В целом естественное плодородие этих почв выше, чем у подзолистых и глее-подзолистых. Они являются преобладающими в пахотном фонде почв таежно-лесной зоны.
Для некоторых почв подзолистого типа большое значение имеет их тепловой режим.
В восточных районах, в условиях континентального климата и недостатка тепла, темные и богатые гумусом дерново-подзолистые и дерново-лесные почвы плохо нагреваются и не обеспечивают корневую систему растений необходимым количеством тепла, что приводит к недозреванию некоторых посевных к/льтур.
Таким образом, наиболее характерными свойствами, присущими почвам дерново-подзолистого типа, являются следующие.
1. Почвы дерново-подзолистого тина не насыщены основаниями, т. е. они содержат в себе то или иное количество обменных ионов водорода и алюминия.
В связи с ненасыщенностью основаниями эти почвы обладают кислыми свойствами.
В результате процессов минерализации органического вещества и выщелачивания эти почвы обеднены гумусом и минеральными легко подвижными соединениями.
Почвы дерново-подзолистого типа заключают в себе сравнительно небольшие количества необходимых для растений питательных веществ.
Эти почвы в большинстве случаев обладают слабо выраженной, непрочной структурой, вследствие чего они способны легко распыляться, заплывать при увлажнении, образовывать корку1 при высыхании и уплотняться. Почвы дерново-подзолистого типа отличаются резко выраженной дифференциацией -почвенного профиля на составляющие их генетические горизонты, причем гумусовый горизонт в большинстве случаен имеет малую мощность.
В составе гумуса видное или первое место принадлежит фульвокислотам, относительное количество которых тем больше, чем резче выражен подзолистый процесс. При нарастании дернового процесса увеличивается количество гуминовых веществ.
Хотя состав почвенных микроорганизмов изучен еще слабо, по все же можно сказать, что в подзолистых почвах ведущее место принадлежит грибам.
Из-за большого разнообразия условий почвообразования и дерново-подзолистой зоне развитие почвообразовательного процесса в почвах может протекать и проявляться самым различным образом, поэтому и отмеченные выше свойства сильно варьируют.
Здесь можно встретить почвы и кислые, и с нейтральной реакцией, и сильно выщелоченные, и почвы с мощным перегнойным горизонтом, и богатые гумусом, в которых подзолообразонательныи процесс совсем почти не выражен, и т. д.
В этом отношении почвенный покров таежно-лесной зоны представляет собой сочетание множества почвенных разновидностей, отличающихся между собой как морфологическими и биохимическими свойствами, так, следовательно, и своим природным плодородием.
При этом, чем сильнее развит дерновый горизонт и чем богаче, следовательно, почва органическим веществом, тем лучше она по своим производственным свойствам, тем выше ее природное плодородие.
На дерново-подзолистой почве можно выращивать практически все культуры, которые хорошо переносят кислую реакцию почву. Так же можно выращивать культуры, которые предпочитают слабокислую и нейтральную реакцию с проведением мелиоративных мероприятий – известкование почвы.
Агрохимическая характеристика дерново-подзолистых почв РФ
Дерново-подзолистые почвы имеют кислую реакцию (pH=4-4,5), значительную обменную кислотность (1-2 м.-экв. на 100 г), 80-90% от которой приходится на обменный Al, а также гидролитическую кислотность (3-6 м.-экв. на 100 г), низкую ёмкость поглощения (5-15 м.-экв.) и степень насыщенности основаниями (30-70%). Большая часть этих почв нуждается в известковании.
Для дерново-подзолистых почв характерно низкое содержание гумуса, общего азота и фосфора и резкое снижение их с глубиной профиля. Агрохимические свойства этих почв сильно варьируют в зависимости от их химического состава и степени окультуренности. В суглинистых почвах содержится от 2 до 3-4% гумуса, 0,1-0,2% азота, 0,07-0,12% фосфора и 1,5-2,5% калия. Валовой состав гумуса (в слое 0-20 см) составляет 60-80 т, азота 3-6 т, фосфора 2-3,5 т и калия 45-75 т на 1 га. Песчаные и супесчаные почвы беднее гумусом, N, P, K, Ca, Mg и микроэлементами, чем суглинистые. Содержание гумуса в лёгких почвах не превышает 0,5-1,0%, азота 0,03-0,08%, фосфора 0,03-0,6% и калия 0,5-1,0%, валовой состав их составляет: 15-30 т, азота 0,9-2,4 т, фосфора 0,9-1,8 т и калия 15-30 т на 1 га. Содержание микроэлементов в дерново-подзолистых почвах колеблется в широких пределах, возможен как недостаток некоторых из них (B, Mo и др.), так и избыток (например, Mn).
Большинство дерново-подзолистых почв характеризуется сравнительно низким содержанием усвояемых (минеральных) форм азота, подвижного фосфора, а песчаные и супесчаные – также и калия. Размер кислотности и содержание подвижных форм питательных веществ в почвах в сильной степени зависит от их окультуренности (табл. 1).
Табл. 1. Агрохимическая характеристика дерново-подзолистых почв
| Степень окультуренности | pH солевой вытяжки | Мощность пахотного горизонта, см | Содержание гумуса, % | Подвижный фосфор | Обменный калий | |
| в мг на 100 г почвы | ||||||
| Слабая | 4-4,5 | до 20 | 1,5-2 | до 5 | до 10 | |
| Средняя | 4,6-5,0 | 20-22 | 2-3,5 | 5-10 | 10-15 | |
| Сильная | 5,1-6,0 | 22-25 | 2,5-4 | 15-25 | 20-30 | |
С повышением степени окультуренности этих почв (при систематическом применении органических и минеральных удобрений, известковании и т.д.) снижается кислотность, увеличивается содержание гумуса и общего азота, подвижного фосфора и обменного калия, повышается их плодородие.
Вследствие того, что дерново-подзолистые почвы обычно бедны элементами питания, но достаточно увлажнены, применение органических и минеральных удобрений даёт высокий эффект. Из минеральных удобрений наиболее эффективны, и их в первую очередь необходимо вносить, азотные, а на слабоокультуренных почвах также фосфорные удобрения. На песчаных и супесчаных почвах эффективно применение калийных, а также магнийсодержащих удобрений.
Биологические особенности культуры
Турнепс – высокоурожайная холодостойкая культура, менее других корнеплодов требовательна к уровню плодородия почвы. Благодаря этому они распространенна до самых северных границ земледелия.
Ботаническое описание
Турнепс (Brassica rapa) – двулетние перекрестноопыляющееся растение семейства Капустные.
Всходы турнепса, при прорастании, выносят на поверхность почвы широкие, с выемкой на конце, семядоли зеленого цвета.
Настоящие листья простые, слабо- и сильнорассеченные. У турнепса листья гладкие, с восковым налетом, опушенные.
Соцветие турнепса – щиток. Цветки с желтоокрашенным четырех-лепестковым венчиком, шестью тычинками и пестиком, развивающимся в многогнездный плод – стручок. У сортов с желтой окраской корнеплода венчик окрашен более интенсивно.
Семена мелкие, коричневые или черные, шаровидные, масса 1000 семян – 2,5-3г.
Биологические особенности
Турнепс холодостоек. Его всходы выдерживают заморозки – до …-5°С, а взрослые растения – до …-6°С. Затяжная холодная весна способствует появлению цветухи. Турнепс умеренно требовательны к теплу, поэтому в южных районах нашей страны они плохо переносят жару и недостаток влаги. Кроме того, здесь их сильнее повреждают насекомые.
Среди корнеплодов турнепс наиболее влаголюбив, поэтому его посевы надо размещать на пониженных местах рельефа, лучше обеспеченных влагой. Повышенная потребность во влаге, в первый год жизни, проявляется у них в начале вегетации. Корневая система турнепса и брюквы распространяется на глубину до 1-1,5м и в ширину – на 40-50см.
Турнепс– растения длинного дня, но требовательны к интенсивности освещения. Турнепс хорошо растет на легких почвах; тяжелые по гранулометрическому составу почвы малопригодны для него. Для этих культур предпочтительна слабокислая реакция почвенного раствора (рНсол=6-6,5), но они удовлетворительно выдерживают повышенную кислотность (рНсол – до 4,3).
С 1т корнеплодов и соответствующим количеством листьев турнепс вносит азота – 4,8кг, фосфора – 1,7кг, калия – 5,7кг.
Период вегетации у турнепса в первый год жизни длится – 70-110 дней, в зависимости от особенности сорта и района возделывания..
Место в севообороте
При выращивании турнепса в овощных севооборотах следует избегать размещения их после капусты и других культур семейства Капустные из-за общих вредителей и болезней. Непригодны также участки, сильно зараженные проволочником. Экономически выгоднее размещать корнеплоды в прифермских кормовых севооборотах.
Обработка почвы
После уборки предшественника, немедленно приступают к лущению стерни. При плотной почве, грубостебельных предшественниках и т.д., проводят 2х кратное лущение дисковыми лущильниками или тяжелыми дисковыми боронами. После прорастания основной массы сорняков, проводят глубокую вспашку плугами с предплужниками на глубину 27-30см. До наступления зимы, зябь обрабатывают по мере необходимости (при появлении сорняков), по типу полупара. Если нет необходимости обработки, то оставляют зябь до весны. Рано весной (при наступлении физической спелости почвы) проводят культивацию с одновременным боронованием, с целью выравнивания почвы и сохранения влаги. За 5-7 дней до посева, проводят предпосевную культивацию с боронованием фрезерными культиваторами КФТ – 3,6 или другими на глубину – 3-4см.
Посев
Турнепс обычно сеют с конца мая и до середины июня для сокращения потерь во время хранения. Турнепс выращивают только посевом семян, поскольку он плохо переносит пересадку.
Норма высева семян турнепса 0,5-0,8 млн/га, в зависимости от засоренности и плодородия почвы.
Требования культуры к элементам питания
Азот является одним из основных элементов питания. турнепс потребляет его в течение всей вегетации, но максимальное количество азота потребляется в первой половине вегетации. Основным источником пополнения почвы азотом служит азот почвы и удобрений. Азот необходим для образования белков. Недостаточное обеспечение сахарной свеклы этим элементом приводит к пожелтению листьев, раннему отмиранию более старых листьев, формированию мелких корней среднего качества.
Фосфор – содержится в растениях в виде сложных белков (нуклеопротеидов), нуклеиновых кислот, фосфатидов, фитина, фосфорных эфиров. Он входит также в состав ферментов и других биологически активных веществ. Значительное количество фосфора находится в растениях в минеральной форме и используется в различных реакциях фосфорилирования.
Калий – необходимый элемент для жизнедеятельности растений. Он обеспечивает течение процесса фотосинтеза, активизирует деятельность многих ферментов. Калий повышает гидрофильность (оводненность) коллоидов протоплазмы клеток. Оптимальное обеспечение растений калием повышает содержание сахара в плодах и овощах. Хорошее калийное питание повышает у растений устойчивость к возбудителям грибковых болезней, к экстремальным погодным явлениям.
Калия особенно много в молодых растениях, в которых энергично делятся клетки. К этому периоду приурочено максимальное поглощение его растениями. В корнеплодах калий накапливается почти до уборки урожая. При недостатке калия пластинки листьев свеклы по краям подсыхают, начиная с наиболее деятельных средних листьев. При этом резко снижается содержание сахара в корнеплодах.
Микроэлементы играют большую роль в жизни растений (натрий, магний, кальций, сера, железо, марганец, бор, медь). Они входят в состав многих ферментов, оказывают влияние на все физиолого-биохимические процессы метаболизма растений, фотосинтез, содержание хлорофилла в листьях, отток ассимилятов, водоудерживающую способность листьев растений и др./6/.
Для формирования урожая корнеплодов турнепс использует значительное количество элементов минерального питания. На образование 1 тонны корнеплода с соответствующим количеством ботвы сахарная свекла потребляет азота 4,8 кг; фосфора – 1,7 кг; калия – 5,7 кг. Коэффициент использования из почвы: азота – 70%; фосфора – 35%; калия – 50%. Коэффициент использования из минеральных удобрений: азота – 65%; фосфора – 135%; калия – 75%.
Особенности микробиологического ценоза и уровень биологической активности
Уровень биологической активности пахотного слоя окультуренной дерново-подзолистой почвы ряда областей был близок к уровню биологической активности выщелоченной черноземной, но ниже, чем в некоторых типах почв Центральных районов Нечерноземной зоны (луговой зернистой, лугово-болотной и др.). Суглинистые почвы более биологически активны, чем супесчаные, особенно по биохимическим показателям. Абиотичные и антропогенные факторы существенно влияют на активность микробоценоза.
В дерново-подзолистой почве численность микроорганизмов находится в следующих пределах: актиномицетов 300-500, грибов 13-182 тыс./г, активность уреазы 95-520 мг N-NH4 в кг почвы, фосфатазы 2,11-6,15 мкг фенолфталеина в 1 г почвы, инвертазы 104-254 мг сахара в 5 г почвы.
Исследования численности микроорганизмов в дерново-подзолистой суглинистой и супесчаной почвах показали, что населенность суглинков значительно плотнее, выше в них более чем на 70% и нитрификационная способность.
Биологическая активность дерново-подзолистой почвы несколько ниже, чем у некоторых других типов почвы Нечерноземной зоны. В этих почвах в большинстве случаев меньше грибов, ниже активность инвертазы, выше нитрификационная способность и активность фосфотазы. Среди дерново-подзолистых почв суглинистые более биологически активны, чем супесчаные. В менее окультуренной супесчаной почве, по сравнению с суглинистой, ниже нитрификационная способность и населенность микроорганизмами, фосформинерализующими, актиномицетами, продуцентами фосфотазы, а в более окультуренной супесчаной почве, главным образом, ниже биохимическая активность (нитрификационная способность, активность инвертазы, фосфатазы). Показатели биологической активности почвы, особенно численность микроорганизмов, под влиянием абиотических и антропогенных факторов очень существенно колеблются и часто соответствуют средним значениям некоторых других типов почв.
Если эти количества микроорганизмов пересчитать, как указывает М. В. Федоров, на живой вес бактериальной массы, приходящейся на пахотный слой гектара почвы, получаются величины следующего порядка: для окультуренных дерново-подзолистых почв — 3,5 т. Распределение бактерий по вертикальному профилю почвы весьма неравномерно: по мере углубления количество микроорганизмов сильно уменьшается. Многие агрономически важные группы микроорганизмов (нитрифицирующие, азотфиксирующие, целлюлозоразлагающие и др.) совсем пропадают или число их так сильно
ограничивается, что они перестают играть существенную роль. В нижних горизонтах дерново-подзолистых почв они, как правило, очень слабо представлены.
Низкая микробиологическая активность нижних горизонтов дерново-подзолистых почв объясняется плохой аэрацией, кислой реакцией,
недостатком питательных веществ. Однако эта особенность, выражающаяся
в уменьшении численности микроорганизмов с глубиной по профилю
почвы, не всегда находит свое проявление в почве.
Исследования показывают, что почти во все сроки наблюдения содержание всех групп сапрофитных микроорганизмов в дерново-подзолистых почвах выше в иллювиальном горизонте по сравнению с вышележащим подзолистым .
Такая аномалия в распределении микроорганизмов по профилю
почвы определяется различием свойств подзолистого и иллювиального
горизонтов и в первую очередь разным содержанием гумуса, являющегося
энергетическим и питательным материалом для сапрофитных микроорганизмов.
При вовлечении целинных почв в пашню численность микроорганизмов в верхнем слое резко увеличивается, а также в какой-то степени и в нижележащих горизонтах почвы. И чем более окультурена почва, тем
больше в ней содержится микроорганизмов, имеющих агрономическое
значение.
Распределение микроорганизмов в почвах разных типов зависит
также от содержания гумуса.
Таким образов в дерново-подзолистой почве содержаться небольшое количество микроорганизмов и в количественном и качественном распределении почвенных микроорганизмов наблюдается определенная зональность. зависящая от экологических условий разных типов почв.
Распределение бактерий по почвенным горизонтам дерново-подзолистой почвы
(в млн. на 1 г почвы) (по С. Разумову и Н. Ремезову)
Генетический горизонт |
Глубина, в см |
Луг | Глубина, в см |
Пашня | Глубина, в см |
Лес |
А1 |
1,0-4,5 | 955,3 | 2-6 | 1086,0 | 0-2,5 | 2693,0 |
| 4,5-7,5 | 852,9 | 10-15 | 982,4 | 2,5-5 | 2246,6 | |
| 7,5-11 | 565,9 | - | - | 5,8 | 1781,5 | |
| А1 А2 | 11-15 | 402,6 | - | - | 9-12 | 782,6 |
| 15-19 | 87,1 | 618,2 | 618,2 | 12-15 | 517,0 | |
| А2 | 19-23 | 71,0 | 382,5 | 382,5 | 16-20 | 355,9 |
| А2 В1 | 23-28 | 50,8 | - | - | 21-25 | 265,6 |
Влияние средств химической защиты и мелиорантов, микроудобрений на биологическую активность и плодородие почвы.
Обработка почвы, особенно вспашка, оказывает определенное влияние на водный, воздушный и тепловой режимы ее. При создании в почве благоприятных условий усиливается развитие микроорганизмов,
способствующих мобилизации питательных веществ, что улучшает условия
питания растений.
Разные приемы обработки почвы по-разному воздействуют на
почвенную микрофлору и мобилизацию питательных веществ в пахотном
слое. Результаты исследований показали, что для активации микробиологических процессов недостаточно одного рыхления почвы. Только рыхление с оборотом пласта способствует значительному увеличению содержания микроорганизмов в пахотном слое почвы. Процессы нитрификации и разложения клетчатки происходят более интенсивно также в случае рыхления и оборота пласта.
При обработке почвы происходит минерализация органического вещества, что приводит к мобилизации питательных веществ. Это показывает, насколько существенное значение имеет обработка почвы для сельскохозяйственного производства и в какой тесной связи с ней находится
активность отдельных групп микроорганизмов, участвующих в мобилизации питательных элементов для растений.
Существенным фактором, определяющим микробиологическую активность почвы, является внесение органических и минеральных удобрений. Органические удобрения как при разовом внесении, так и при длительном применении всегда оказывают положительное влияние на развитие микроорганизмов в почве и на их биохимическую активность.
Систематическое внесение навоза в почву значительно повышает
общее количество микроорганизмов в ней, при этом увеличивается число не
только бактерий, но и актиномицетов и грибов. Внесение в почву
минеральных удобрений также благоприятно влияет на развитие почвенных
микроорганизмов. Минеральные удобрения стимулируют развитие
микроорганизмов, что ускоряет процессы разложения органического
вещества почвы. Необходимо, однако, отметить, что в тех случаях, когда в
почве мало свежих органических соединений и перегноя, то длительное
внесение минеральных удобрений приводит к постепенному уменьшению
числа микроорганизмов.
Длительное применение минеральных удобрений в севообороте, особенно при монокультуре, приводит к резкому падению численности бактерий в почве. Причинами такого явления следует считать изменение активной реакции почвенного раствора, появление почвенного токсикоза, недостаток в энергетическом материале и односторонность в питательном режиме почвы. Длительное же применение минеральных удобрений в сочетании с органическими удобрениями обеспечивает высокую биологическую активность почвы.
Важным фактором, определяющим состав и численность микрофлоры почвы и характер микробиологических процессов, является реакция
почвенной среды. Почвы как кислые, так и имеющие сильнощелочную
реакцию неблагоприятны и для микрофлоры, и для высших зеленых
растений. Известкование резко увеличивает численность бактерий в почве.
Для уменьшения щелочности почвы может быть применена сульфофикация (внесение в почву элементарной серы). Этот прием хотя и дает хорошие результаты, но еще не имеет широкого распространения из-за
дороговизны.
Общее количество аэробных бактерий в дерново-подзолистой почве
(в млн. на 1 г почвы)
| Вариант | Сроки наблюдений | ||
| В севообороте | |||
| Без удобрений | 5,8 | 3,4 | 5,9 |
| Без удобрений + известь | 20,7 | 6,9 | 14,0 |
| Навоз + РК | 52,6 | 51,7 | 68,9 |
| Навоз | 54,9 | 31,5 | 37,3 |
| Навоз + РК + известь | 149,5 | 54,9 | 138,5 |
| РК + известь | 32,8 | 15,1 | 22,3 |
| РК | 19,7 | 9,7 | 14,9 |
| N | 15,6 | 12,8 | 8,4 |
| В монокультуре | |||
| Без удобрений | 3,3 | - | 4,6 |
| Без удобрений + известь | 17,3 | 2,2 | 11,4 |
| Навоз + РК | 45,4 | 19,6 | 30,6 |
| Навоз + РК + известь | 92,1 | 30,0 | 65,8 |
| РК + известь | 13,9 | 8,1 | 12,6 |
| РК | 2,3 | 5,7 | 4,9 |
| N | 7,1 | 1,2 | 4,6 |
Почвы как кислые, так и имеющие сильнощелочную реакцию неблагоприятны и для микрофлоры, и для высших зеленых растений. Известкование резко увеличивает численность бактерий в почве.
Велика роль почвенных микроорганизмов при синтезе и распаде
гумуса почвы, в создании почвенной структуры и других почвенных
процессах.
Изменение численности микрофлоры в почве в связи с применением удобрений
(количество микроорганизмов в тыс. на 1 г почвы)
| Варианты | Общее количество микроорганизмов |
бактерии | Актиномицеты | Плесневые грибы | ||||
| до | после | до | после | до | после | до | после | |
| Без удобрений | 1180 | 538 | 470 | 350 | 700 | 180 | 10 | 8 |
| Навоз | 2550 | 1135 | 830 | 610 | 1700 | 510 | 20 | 16 |
| Навоз + известь | 1983 | 1397 | 760 | 530 | 1200 | 850 | 23 | 17 |
| Навоз + РК | 1101 | 1205 | 366 | 540 | 700 | 640 | 35 | 25 |
| Показатели | Удобрения | ||
| N | P | K | |
| Вынос питательных веществ на 1 т основной продукции, кг | 4.8 | 1.7 | 5.7 |
| Вынос питательных веществ с планируемым урожаем | 960 | 340 | 1140 |
| Содержание подвижных форм питательных веществ, мг/кг | 23 | 22 | 36 |
| Содержание доступных форм питательных веществ кг/га | 690 | 660 | 1080 |
| Коэффициент использования питательных веществ из почвы кг/га | 35 |
15 |
30 |
| Использование питательных веществ из почвы кг/га | 241.5 | 99 | 324 |
| Будет внесено с 60 т навоза | 300 | 150 | 330 |
| Коэффициент использования питательных веществ из органических удобрений кг/га | 53 |
45 |
68 |
| Будет усвоено из органических удобрений | 159 | 67.5 | 224.4 |
| Будет усвоено из почвы | 400.5 | 166.5 | 548.4 |
| Требуется внести с минеральными удобрениями | 559.5 | 173.5 | 591.6 |
| Коэффициент использования питательных веществ из минеральных удобрений кг/га | 65 |
35 |
75 |
| Требуется внести с минеральными удобрениями с учетом коэффициента использования | 363.7 |
60.7 |
443.7 |
| N 363.7 P 60. 4 K443.7 | |||
| Потребность внесения удобрения к физической массе минерального удобрения кг/га | |||
| Мочевина | 46,2 | 7.8 | |
| Суперфосфат двойной | 26 | 2.3 | |
| Хлористый калий | 62,5 | 7.0 | |
Оптимальная доза внесения по турнепс N140 P60. K150
Расчет нормы внесения удобрений
| N | P | K | |
| Предпосевная обработка (ранневесеннее внесения) + навоз | 0,1 | 378,3 | 00,6 |
| Основная обработка (при вспашке) | 226,5 | 756,6 | 616,1 |
| Подкормка | 113,2 | 0,1 | 308,0 |
Известкование почвы
Доза извести CaCO3 = 6 т.га
Сроки внесения удобрений могут быть различные: это допосевное (основное), припосевное (или припосадочное) и послепосевное (подкормка) удобрения.
Допосевное удобрение включает всю годовую норму органических удобрений и наибольшую часть минеральных. Это удобрение заделывают в более глубокие увлажненные слои почвы, откуда его могут использовать растения в течение всего вегетационного периода.
Припосевное удобрение вносят одновременно с посевом (посадкой) сельскохозяйственных культур на глубину 2-3см глубже семян (клубней). Это удобрение обеспечивает растения легкоусвояемыми питательными веществами, особенно фосфором, в самый начальный период жизни, когда они имеют слаборазвитую корневую систему. Нормы припосевного удобрения значительно меньше норм основного, так как растения используют его только в первые 2-3 недели жизни.
Послепосевное удобрение усиливает питание растений в отдельные критические периоды их развития, когда необходимы те или иные элементы.
Способы внесения удобрений и его сроки определяются почвенно-климатическими условиями, особенностями возделываемых культур, свойствами удобрений и организационно-хозяйственными возможностями.
Одноразовое внесение удобрений до посева оправдывается только в том случае, если применяют достаточное количество удобрений в требуемом соотношении питательных веществ. При этом снижаются затраты на удобрения и повышается их окупаемость. Многочисленными исследованиями установлено, что наиболее высокая прибавка урожая яровых культур достигается при заделывании допосевного удобрения осенью под зяблевую вспашку. Однако на легких супесчаных и песчаных почвах, на почвах с близким залеганием грунтовых вод, на крутых склонах, в районах с избыточным увлажнением, где возможны потери внесенных с осени удобрений путём их вымывания или смыва, основное удобрение следует давать в весенний период.
При посеве зерновых культур используют, как правило, гранулированный суперфосфат. 1ц простого гранулированного суперфосфата при рядковом внесении даёт дополнительно 5-6ц зерна, а при разбросном - только 1-2ц. При рядковом способе гранулированный суперфосфат используют в небольшом количестве – 10-15кг/га. Этой нормы вполне достаточно для нормального развития всходов в первый период их роста.
Весьма эффективным приёмом для повышения качества зерна являются некорневые подкормки в более поздние сроки. Нормы удобрений при некорневых подкормках устанавливают по результатам диагностики питания растений в период их вегетации.
Для кукурузы наиболее эффективна ранняя подкормка при первой междурядной обработке.
Удобрения вносят вразброс с последующей заделкой в почву или локально. При разбросном внесении удобрений должно быть обеспечено сплошное равномерное распределение удобрений по поверхности почвы. При локальном внесении туки размещаются в почве очагами различной формы. Локальное внесение удобрений по сравнению с разбросным повышает их эффективность до 20% и более. Оптимальная глубина размещения лент удобрения – 10см на суглинистых и глинистых почвах, 15см – на супесчаных и песчаных почвах при ширине лент 2-4см и расстоянии между лентами 16-20см.
Рациональное применение удобрений должно быть обеспечено только при внедрении научно обоснованной системы удобрения в севообороте. При возделывании сельскохозяйственных культур в севообороте создаются лучшие условия для более эффективного применения удобрений в связи с меньшей засоренностью посевов, более слабыми повреждениями растений специфическими вредителями и болезнями, более благоприятным водным режимом. Эффективность применения удобрений в севообороте повышается на 20-30%. При разработке системы удобрения необходимо учитывать особенности севооборота, почвенно-климатические и агротехнические условия.
- Аринушкина Е.В. Руководство по химическому анализу почв. М.: Изд-во МГУ, 1970. 487 с.
- Благодатская Е.В., Пампура Т.В., Мякшина Т.Н., Демьянова Е.Г. Влияние растворимых и нерастворимых соединений свинца на дыхательную активность микроорганизмов серой лесной почвы в многолетнем полевом эксперименте // Почвоведение. 2006. № 5. С. 559–568.
- Благодатский С.А., Благодатская Е.В., Горбенко А.Ю., Паников Н.С. Регидратационный метод определения микробной биомассы в почве // Почвоведение. 1987. № 4. С. 64–71.
- Евдокимов И.В., Саха С., Благодатский С.А., Кудеяров В.Н. Иммобилизация азота почвенными микроорганизмами в зависимости от доз внесения N // Почвоведение. 2005. № 5. С. 581–89.
- Евдокимова Г.А., Зенкова И.В. Влияние выбросов алюминиевого завода на биоту почв Кольского полуострова // Почвоведение. 2003. № 8. С. 973–979.
- Левин СВ., Гузев B .C ., Асеева И.В. и др. Тяжелые металлы как фактор антропогенного воздействия на почвенную микробиоту // Микроорганизмы и охрана почв. М.: Изд-во МГУ, 1989. С. 5–46.
- Лукина Н.В., Никонов В.В., Базель B .C . Рассеянные элементы в бореальных лесах / Отв. ред. Исаев А.С. М.: Наука, 2004. 616 с.
- Марфенина О.Е. Реакция комплекса микроскопических грибов на загрязнение почв тяжелыми металлами // Вестник МГУ. Сер. 17, почвоведение. 1985. № 2. С. 46–50.
- Никонов В.В., Лукина Н.В., Полянская Л.М.и др. Численность и биомасса почвенных микроорганизмов северо-таежных сосновых лесов при пирогенной сукцесоии // Почвоведение. 2006. № 4. С. 484–494.
- Паников Н.С. Кинетика роста микроорганизмов: общие закономерности и экологические приложения. М.: Наука, 1992. 311 с.
- Полянская Л.М., Никонов В.В., Лукина Н.В. и др. Микроорганизмы Al-Fe-гумусовых подзолов сосняков лишайниковых в условиях аэротехногенного загрязнения // Почвоведение. 2001. № 2. С. 215–226.
- Смирнова О.В., Бобровский М.В. Онтогенез дерева и его отражение в структуре и динамике растительного и почвенного покрова // Экология. 2001. № 3. С. 177–181.