Состав мембран
СОДЕРЖАНИЕ: Белки и липиды как основные компоненты мембран. Фосфолипидный состав субклеточных мембран печени крысы. Длинные углеводородные цепи. Мембраны грамположительных бактерий. Пути биосинтеза мембранных липидов и механизмы их доставки к местам назначения.Основными компонентами мембран являются белки и липиды. На долю углеводов может приходиться около 10% массы мембран, при этом они всегда входят в состав гликолипидов или гликопротеи-нов. Соотношение между белками и липидами в мембранах значительно варьирует — от 20% белка в миелине до 80% в митохондриях. В табл. 1.3 и 1.4 обобщены данные по составу ряда мембран. Плотность мембран прямо пропорциональна содержанию в них белка. Судя по данным изопикнического центрифугирования, чем выше содержание белка в мембране, тем больше ее плотность.
Таблиця 1. Фосфолипидный состав субклеточных мембран печени крысы
| Доля от суммарного количества фосфолипидов, It | ||||||
| митохондрии | микро-сомы | ли10- сомы |
плазматическая мембрана | ядерная мембрана | мембраны аппарата Гольджи | |
| Кардиолипин | 18 | 1 | 1 | 1 | 4 | 1 |
| Фосфатидилэтаноламин | 35 | 22 | 14 | 23 | 13 | 20 |
| Фосфатидилхолин | 40 | 58 | 40 | 39 | 55 | 50 |
| Фосфатидилииозитол | 5 | 10 | 5 | 8 | 10 | 12 |
| Фосфатидилсерин | 1 | 2 | 2 | 9 | 3 | 6 |
| Фосфатидная кислота | — | 1 | 1 | 1 | 1 | |
| Лиэофосфоглицериды 2 | 1 4 | 11 | 7 | 2 | 3 | 3 |
| Сфингомиелин | 1 | 1 | 20 | 16 | 3 | 8 |
| Фосфолипиды | 0,175 | 0,374 | 0,156 | 0,672 | 0,500 | 0,825 |
| Холестерол | 0,003 | 0,014 | 0,038 | 0,128 | 0,038 | 0,078 |
Таблица 2 Белковый и липидиый состав некоторых мембран животных и бактериальных клеток. Л/Б — отношение липка/белок

Белковый состав мембраны зависит в какой-то степени от метода ее выделения. Некоторые белки непрочно связаны с мембраной и легко удаляются при промывании ее растворами с высокой или, напротив, с низкой ионной силой, щелочными растворами или растворами, содержащими хелатирующие агенты типа ЭДТА. Бывают и случаи, когда трудно сказать, является ли белок мембранным или цитоплазматическим, случайно связавшимся с мембраной в ходе ее выделения.
1. МЕМБРАННЫЕ ЛИПИДЫ
Наиболее поражает в мембранных липидах их огромное разнообразие. Причины этого пока не ясны, хотя становится все более очевидно, что, по-видимому, связано это с тем разнообразием функций, которые липиды выполняют в мембранах. Но, конечно, главная функция мембранных липидов состоит в том, что они формируют бислойный матрикс, с которым взаимодействуют белки. Основные классы липидов представлены на рис. 1.8; их краткое описание дано ниже.
Глицерофосфолипиды
Это наиболее распространенные липиды. Одна из гидроксильных групп глицерола связана с полярной группировкой, содержащей фосфат, а две другие — с гидрофобными остатками. Номенклатура гли-церидов основана на системе стереоспецифической нумерации. Если глицерол изобразить в проекции Фишера, так что центральная группа будет расположена слева, то атомы углерода будут нумероваться так, как показано на рис. В этом случае в название глицерофосфолипида вводят приставку snдля обозначения положения заместителя. В литературе встречается несколько систем стереохимических обозначений: sn, D/L и R/S. На рис. представлена стереохимия атома С-2 в этих трех системах. Природные фосфолипиды.
У большинства фосфоглицеридов фосфатная группа находится в ял-3-положении глицерола; она обычно связана с какой-либо из групп, представленных на рис.
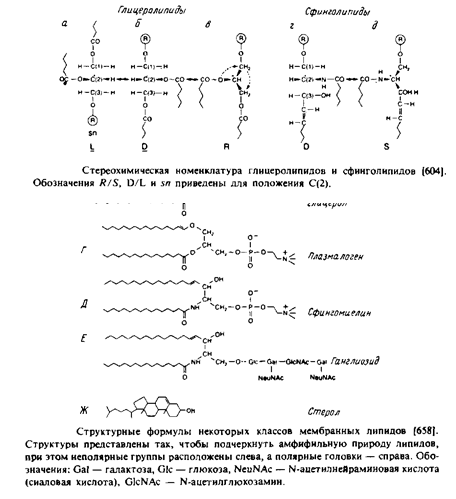
Длинные углеводородные цепи, находящиеся в положениях sn-\ и sn-2, могут присоединяться за счет сложноэфирной и простой эфирной связей. Эти цепи значительно различаются по длине, раз-ветвленности и степени ненасыщенности.
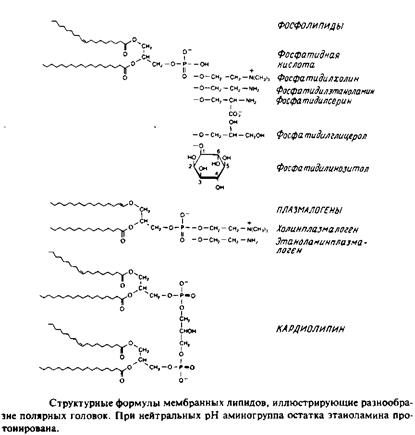
1. 1,2-диацилфосфоглицериды или фосфолипиды. Эти липиды, являющиеся сложными эфирами жирных кислот и глицерола, широко представлены во многих мембранах эукариотических и прокарио-тических клеток, за исключением архебактерий. Фосфатидил-холин является основным компонентом мембран животных клеток, а фосфатидилэтаноламин — это нередко основной липид бактериальных мембран. В табл. представлен ряд жирных кислот, наиболее часто встречающихся в составе фосфолипидов, а в табл. приведен жирнокислотный состав клеточных мембран печени крысы.

Жирные кислоты почти всегда содержат четное число атомов углерода в пределах от 14 до 24. Наиболее распространены кислоты С16, С18 и С20. Степень ненасыщенности может быть разной, но чаще всего встречаются ненасыщенные кислоты 18:1, 18:2, 18:3 и 20:4. Здесь первое число обозначает длину ацильной цепи, а второе указывает на число содержащихся в ней двойных связей. Почти все природные кислоты характеризуются г/ис-конфигурацией двойных связей. Цепь в такой конфигурации имеет излом, что нарушает упаковку липидных молекул в бислое. В составе молекул многих фосфолипидов имеется одна насыщенная и одна ненасыщенная цепи. В случае животных клеток ненасыщенные цепи обычно находятся в 5и-2-положении глицерола. Такое же распределение цепей характерно и для фосфолипидов клеток Е. coli. Двойные связи в полиненасыщенных цепях обычно являются несопряженными. В фосфолипидах некоторых бактериальных мембран обнаружены разветвленные цепи, а также цепи, содержащие циклы, и гидроксильные группы в /3-положении. На рис. 1.11 показана структура некоторых из этих кислот.
2.У архебактерий глицерофосфолипиды имеют обращенную сте-реохимическую конфигурацию, при которой фосфорильные группы находятся в sn-\-положении глицерола. У многих бактерий этого вида гидрофобные компоненты представляют собой не сложные эфиры жирных кислот, а простые изопранильные эфиры глицерола.
Кардиолипины или дифосфатидилглицеролы. По сути это димерные формы фосфолипидов. Они содержатся в боль шом количестве во внутренней мембране митохондрий, в мембране хлоропластов и в некоторых бактериальных мембранах, но редко встречаются в других мембранах.

4. Плазмалогены. Это фосфоглицеролипиды, у которых одна из углеводородных цепей представляет собой простой виниловый эфир. Этаноламиновые плазмалогены широко представлены в миелине и в саркоплазматическом ретикулуме сердца.
Фосфосфинголипиды
Эти липиды имеют такие же полярные головки, как и глицерофосфолипиды, но их гидрофобная часть представлена церамидом. В плазматических мембранах животных клеток широко распространен сфингомиелин. Основными жирнокислотными компонентами в миелине являются кислоты 24:1 и 24:0. В мембранах растительных и бактериальных клеток фосфосфинголипиды встречаются редко. Кроме сфингомиелина известны и другие фосфосфинголипиды, например церамид-1-фосфорилэтаноламин, церамид-1-фосфорилинози-тол и церамид-1-фосфорилглицерол.
Гликоглицеролипиды
Это полярные липиды, у которых в м-З-положении глицерола находится углевод, присоединенный с помощью гликозидной связи, например галактоза. Гликоглицеролипиды широко представлены в мембранах хлоропластов, они обнаружены также в заметных количествах в сине-зеленых водорослях и бактериях. Моногалактозилдиа-цилглицерол был назван «наиболее распространенным в природе полярным липидом», поскольку на его долю приходится половина всех липидов тилакоидной мембраны хлоропластов. Для мембран грамположительных бактерий характерны гликоглице-
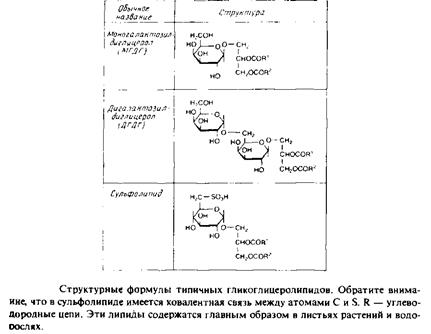
ролипиды с большим разнообразием Сахаров. Архебактерии также содержат такие липиды, но, как и в случае глицерофосфолипидов, их стереохимическая конфигурация является обращенной, с локализацией гликозидной связи в sn-l-положении глицерола. В мембранах животных клеток гликоглицеролипиды встречаются редко.

Гликосфинголипиды
классифицируют в соответствии с размером углеводной части, которая может быть представлена всего лишь одним моносахаридным остатком, с одной стороны, и очень сложным углеводным полимером — с другой. Моногликозилцерамиды обычно называют цереброзидами. Ганглиозиды представляют собой класс анионных гликосфинголипидов, которые содержат один или несколько остатков сиаловой кислоты, связанных с сахарными остатками церамидолигосахарида. Глобозидами называют нейтральные гликосфинголипиды, которые не содержат остатков отрицательно заряженной сиаловой кислоты.
Гликосфинголипиды находятся на наружной поверхности плазматических мембран животных клеток; обычно они являются минорными компонентами, но иногда содержатся в значительных количествах. Моногалактозилцерамид — это один из основных компонентов миелиновой оболочки нервного волокна. В некоторых случаях гликосфинголипиды локализуются не в плазматической мембране, а во внутриклеточных мембранах.
Гликосфинголипиды мембран эритроцитов несут антигены группы крови. В клетках аденокарциномы человека накапливаются необычные фукозилированные гликосфинголипиды, которые можно использовать для обнаружения этих клеток и контроля за развитием опухоли.
Стеролы
Эти липиды присутствуют во многих мембранах растений, животных и микробов. По-видимому, самым распространенным из сте-ролов является холестерол. Его молекула состоит из компактного, жесткого гидрофобного ядра, а полярной головкой является гидро-ксильная группа. Холестерол содержится в плазматических мембранах животных клеток, в лизосомах, эндосомах и в мембранах аппарата Гольджи. Он составляет около 30% всей массы мембранных липидов во многих плазматических мембранах животных клеток. В высших растениях обнаружены другие стеролы, чаще всего ситостерол и стигмастерол. Растительные стеролы часто имеют еще одну боковую цепь в положении С-24 и/или двойную связь в положении С-22. В мембранах дрожжей и дру-
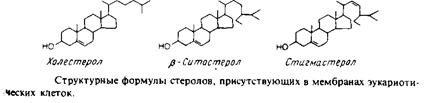
гих эукариотических микроорганизмов часто содержится эргостерол. К классу стеролоподобных липидов относят также гопанои-ды, которые найдены в бактериях и некоторых растениях.
Минорные компоненты
В мембранах присутствуют также и другие липиды, которые можно отнести к разряду минорных компонентов вследствие их малого содержания в мембранах. Так, в мембранах обычно обнаруживаются, хотя и в очень малых количествах, свободные жирные кислоты и лизофосфолипиды. Пожалуй, исключением из этого правила являются мембраны хромаффинных гранул, которые, как известно, содержат необычно много свободных жирных кислот. Минорными компонентами мембран являются также моноацил- и диацилглице-ролы. Диацилглицеролы выполняют важную функцию вторых посредников в передаче сигнала при активации клеток рядом биологически активных веществ. Эта система клеточного отклика на внешний стимул будет детально рассмотрена в гл. 9. В мембранах обычно присутствуют и полиизопреноидные липиды. К ним относятся уби-хиноны и менахиноны — компоненты цепи электронного транспорта в мембранах. Можно отметить также ундекапренол и долихол, которые являются липидными переносчиками промежуточных продуктов соответственно при биосинтезе клеточной стенки у прокариот и при биосинтезе гликопротеинов в аппарате Гольджи эукариот. Длина молекул этих липидов в вытянутом состоянии значительно превышает толщину бислоя, поэтому неизвестно, как эти молекулы в нем расположены. Неясно также, почему липидными переносчиками служат именно полиизопреноидные структуры.
2. МНОГООБРАЗИЕ ФУНКЦИЙ МЕМБРАННЫХ ЛИПИДОВ
Совершенно очевидно, что липидный состав различных мембран не является случайным, однако удовлетворительного объяснения этому феномену не найдено. Любая конкретная мембрана может содержать более ста разных типов липидных молекул. Почему их так много и почему каждая мембрана имеет уникальный липидный состав? Пути биосинтеза мембранных липидов и механизмы их доставки к местам назначения обсуждаются в разд. 10.4. Становится все более очевидным, что липиды активно участвуют в процессах, протекающих в мембранах, однако причины их разнообразия неясны. Рассмотрим некоторые факторы, возможно, определяющие липидный состав мембраны.
1. Смесь липидов обязательно должна быть способна образовать стабильный бислой, в котором могли бы функционировать белки.
2.Некоторые липиды способствуют стабилизации сильно искривленных участков мембраны, образованию контакта между мембранами или связыванию определенных белков, поскольку форма этих молекул благоприятствует нужной упаковке бислоя на соответствующих участках мембраны.
3.Некоторые липиды являются важными биорегуляторами. Наиболее изучена в этом отношении регуляторная роль производных фосфатидилинозитола в плазматических мембранах клеток эукариот.
4.Некоторые липиды участвуют в реакциях биосинтеза. Например, в клетках Е. coliфосфатидилглицерол поставляет глицерофосфатный фрагмент при биосинтезе периплазматических олигосахари-дов.
5.Отдельные липиды необходимы для поддержания оптимальной активности ряда ферментов. Этот вопрос рассматривается в гл. 6.
6.Ганглиозиды, как полагают, играют важную роль в регуляции роста клеток, являются специфическими рецепторами в плазматической мембране и ответственны за клеточную адгезию.
Как было показано экспериментально, организмы часто могут выдерживать — причем без всяких последствий — существенные изменения липидного состава мембран. Например, с помощью генетической трансформации можно получить штаммы Е. coli, в мембранах которых содержится 34% фосфатидной кислоты, обычно отсутствующей в штаммах дикого типа. Очевидно, тот липидный состав, который характерен для штаммов дикого типа, не является обязательным для выживания клеток, по крайней мере, в условиях их выращивания в лаборатории.
3. МЕМБРАННЫЕ БЕЛКИ
Как видно из табл. 1.3 и 1.4, мембраны содержат от 20 до 80% белка. Как правило, именно белки ответственны за функциональную активность мембран. К ним относятся разнообразные ферменты, транспортные белки, рецепторы, каналы, поры и т. д., которые обеспечивают уникальность функций каждой мембраны. Первые успехи в изучении мембранных белков были достигнуты тогда, когда биохимики научились использовать детергенты для выделения мембранных белков в функционально активной форме. Это были работы по изучению ферментных комплексов внутренней мембраны митохондрий. Значительным шагом вперед было осознание того, что мембранные белки имеют не исключительно /3-складчатую структуру, как предполагалось в модели «элементарной мембраны» Дэвсона—Даниелли—Робертсона, а содержат достаточно много а-спиралей. Важное значение имел также вывод о том, что мембранные белки могут глубоко проникать в липидный бислой или даже пронизывать его и что их стабилизация осуществляется за счет гидрофобных взаимодействий. Эти термодинамические представления существенно обогатили принцип «гидрофобных сил», предложенный для объяснения структуры белков и предполагавший существование неполярной, гидрофобной области внутри белковой глобулы и полярных, гидрофильных участков, контактирующих с водной средой.
По мере совершенствования методов очистки удавалось получать в изолированном виде все большее число мембранных белков. Определение первичной структуры большинства из них было затруднено из-за плохой растворимости в воде как самих белков, так и получаемых из них гидрофобных пептидов. В середине 1970-х гг. эта проблема была решена для двух мембранных белков — гликофорина и цитохрома bs, что позволило установить основной принцип структурной организации интегральных белков. В аминокислотной последовательности гликофорина — сиалогликопротеина из мембраны эритроцитов — был обнаружен короткий участок, состоящий из 23 неполярных аминокислот и расположенный примерно в середине цепи. Данные топологических и других исследований показали, что молекула гликофорина полностью пронизывает мембрану, причем погруженный в мембрану гидрофобный участок имеет а-спиральную конфигурацию. Так вошла в жизнь новая, теперь уже общепризнанная концепция о наличии в мембранных белках а-спиральных доменов, пронизывающих мембрану. Эта концепция была полностью подтверждена при изучении трансмембранных белков с помощью методов, которые позволяют получить максимально возможное в наше время разрешение. Судя по результатам реконструкции электронно-микроскопических изображений препаратов бактериородопсина из пурпурной мембраны Halobacteriumhalobiumи по данным рентгеноструктурного исследования фотосинтетических реакционных центров бактерий, эти белки содержат несколько а-спиральных участков, последовательно пересекающих бнслой.
Другой вариант расположения полипептидной цепи в мембране был обнаружен при изучении аминокислотной последовательности интактной формы микросомного цитохрома bi. Было показано, что этот белок содержит относительно короткий участок вблизи карбоксильного конца, состоящий из гидрофобных аминокислот. Этот «гидрофобный якорь» можно было удалить с помощью протеолиза, причем гемсвязывающий домен высвобождался в водорастворимой форме. Локализованный в мембране гидрофобный домен, или «якорь», стал еще одним характерным элементом структуры мембранных белков.
В основе современных представлений о структуре мембранных белков лежит идея о том, что их полипептидная цепь уложена так, чтобы образовалась неполярная, гидрофобная поверхность, контактирующая с неполярной областью липидного бислоя. Полярные или заряженные домены белковой молекулы могут вазимодействовать с полярными головками липидов на поверхности бислоя. Многие мембранные белки являются трансмембранными и пронизывают бислой. Некоторые белки, по-видимому, связаны с мембраной лишь за счет их взаимодействия с другими белками.
Мембранные белки обычно связываются с мембраной с помощью нековалентных взаимодействий — гидрофобных или электростатических сил. Однако есть мембранные белки, которые связаны с липидами ковалентно. Такие примеры пока немногочисленны, но их появляется все больше. Многие белки плазматических мембран растительных и животных клеток относятся к классу гликопротеинов. Углеводные остатки этих белков всегда находятся с наружной стороны плазматической мембраны.
Обычно мембранные белки подразделяют на наружные и внутренние. При этом критерием служит степень жесткости обработки, необходимой для извлечения этих белков из мембраны. Периферические белки высвобождаются при промывании мембран буферными растворами с низкой ионной силой, буферными растворами с низким или, наоборот, высоким значением рН и в присутствии хелатирующих агентов, связывающих двухвалентные катионы. Как полагают, такие белки связаны с поверхностью мембраны за счет слабых электростатических взаимодействий с полярными головками липидных молекул либо с молекулами других белков. Часто бывает нелегко отличить периферические мембранные белки от белков, связавшихся с мембраной в процессе ее выделения. При обработке мембранного препарата буфером с низкой ионной силой в раствор переходит около 30% белков, связанных с мембраной эритроцитов. При несколько более жесткой обработке хаотропными агентами высвобождаются периферические белки. В ряде случаев эти агенты оказывают влияние достаточно сильное, чтобы разрушить белок-белковые взаимодействия, хотя денатурации белков при этом не происходит. Действие хаотропных агентов, «нарушающих структуру воды», обусловлено главным образом ослаблением гидрофобных взаимодействий между компонентами мембраны.
Для высвобождения интегральных мембранных белков необходимо использовать детергенты или даже органические растворители. Детергенты разрушают липидный бислой и, как полагают, связываются с гидрофобными участками мембранных белков, контактирующими с гидрофобной областью бислоя. Для того чтобы сохранить интегральные мембранные белки в растворенном монодисперсном состоянии, в растворе постоянно должны присутствовать детергенты. При удалении детергентов неизбежно происходят агрегация белков и их последующее осаждение. Дальнейшая информация о взаимодействиях между белками и детергентами, а также о структуре и свойствах мембранных белков содержится в гл. 3.
Резюме
Методы дифракции рентгеновских лучей и электронной микроскопии сыграли историческую роль в развитии мембранологии и внесли решающий вклад в современные представления о биологических мембранах. Сегодня не вызывает сомнений, что липидный бислой образует структурную основу практически всех биологических мембран и что их функциональное многообразие основано именно на этом структурном единстве различных мембранных систем. Каждая отдельно взятая мембрана содержит большое число различающихся по своим химическим свойствам липидов. Причины этого разнообразия неясны, хотя появляется все больше данных об уникальных биологических функциях отдельных липидов. На долю белков приходится от 20 до 80% массы мембран. Многие из этих белков полностью пронизывают липидный бислой, и их удается солюбилизировать только с помощью детергентов. Другие мембранные белки, называемые периферическими, легкое извлекаются из мембраны с помощью буферных растворов с изменяющимися рН или ионной силой либо при удалении двухвалентных катионов с помощью хелатирующих агентов.
В настоящее время разработаны методы выделения и характеристики индивидуальных мембран из клеток прокариот, из животных и в меньшей степени из растительных клеток. Разделение чаще всего основано на различиях в размере и плотности мембранных частиц, содержащихся в гомогенате разрушенных клеток. Можно также использоать различия в поверхностных свойствах мембран или в их электрофоретическом поведении. Выделение и очистка мембран — первый и обязательный этап в их биохимическом исследовании.