Структурно-функциональные параллели в организации кортикального ядра миндалевидного комплекса
СОДЕРЖАНИЕ: Уникальность топографии и структурной организации кортикального ядра лежит в основе часто возникающих в морфологической литературе различного рода разногласий: каковы границы этого образования в пределах МК, ядро ли это или корковая формация?Карпова А.В., Бикбаев А.Ф., Калимуллина Л.Б.
Миндалевидный комплекс (МК, corpus amygdaloideum) является ядерно-палеокортикальным компонентом мозга [1]. Его кортикальное ядро, характеризующееся значительной площадью и рострокаудальной протяженностью, располагается на вентральной поверхности мозга и является одной из основных морфообразующих структур. Уникальность топографии и структурной организации кортикального ядра лежит в основе часто возникающих в морфологической литературе различного рода разногласий: каковы границы этого образования в пределах МК, ядро ли это или корковая формация? Важным в данном направлении явилось предпринятое Scalia и Winans [2], Krettek и Price [3] деление кортикального ядра на переднее кортикальное ядро (СОа), периамигдалярную кору (РАС) и заднее кортикальное ядро (СОр). Дифференциация этих структур означала признание проблемы существования ядерно-палеокортикальных взаимоотношений в миндалевидном комплексе.
Между тем в физиологии кортикальное ядро часто рассматривается как единое образование, несмотря на сложность и гетерогенность его структурной организации. Это может являться одной из причин противоречивости результатов исследований функционального значения этой структуры. Поскольку известно, что миндалевидный комплекс играет важную роль в патогенезе височной (лимбической) эпилепсии [4, 5, 6, 7, 8], особую важность приобретает учет структурных особенностей входящих в его состав отделов.
В настоящей работе мы попытались связать особенности структурной организации отделов кортикального ядра с характером их электрической активности.
Материал и методика
Работа выполнена на 65 половозрелых самцах крыс линии Вистар. Цитоархитектоническое исследование проводили по препаратам, окрашенным крезилом фиолетовым по методу Ниссля. Нейронную организацию исследовали по препаратам, импрегнированным нитратом серебра по Гольджи. Использовали классификации Т.А.Леонтович [9] для подкорковых и Г.И.Полякова [10] для корковых нейронов. Для количественной характеристики дендритного дерева нейронов мы впервые применили методы многомерного анализа (корреляционный, факторный и кластерный анализы) с целью получения формального описания нейронов и выявления особенностей структурной организации отделов кортикального ядра.
Изучение электрофизиологических характеристик осуществляли методом электроэнцефалографии. Фоновую ЭЭГ регистрировали посредством хронических нихромовых электродов, вживленных в СОа, РАС, СОр по стереотаксическим координатам [11]. Статистическую обработку и анализ данных (вычисление описательных статистических характеристик, частотно-спектральный анализ ЭЭГ, вычисление относительной спектральной плотности, дисперсионный анализ) производили в программном пакете “STATISTICA 5.1”.
Результаты исследования
Сравнительный анализ цитархитектоники отделов, а также поуровневое исследование нейронной организации кортикального ядра показали, что оно представляет собой гетерогенное образование. На основании результатов анализа, опираясь на учение А.А.Заварзина о ядерных и экранных центрах нервной системы [12] и руководствуясь критериями “коры”, разработанными Pigache [13], мы считаем целесообразным классифицировать отделы кортикального ядра следующим образом:
переднее кортикальное ядро – межуточная формация;
медиальная часть заднего кортикального ядра – межуточная формация;
латеральная часть заднего кортикального ядра – межуточная формация;
заднее кортикальное ядро переходной к гиппокампу зоны – межуточная формация;
периамигдалярная кора рострального уровня центрального отдела – палеокортекс с элементами упрощенности;
периамигдалярная кора каудального уровня центрального отдела и заднего отдела миндалевидного комплекса – палеокортекс.
На основании количественных характеристик нейронов нами показана представленность типов нейронов по отделам кортикального ядра: редковетвистые короткодендритные нейроны преимущественно представлены в СОа (48,0 % от общего числа клеток этого типа), крупные корковые (46,7 %) и малодифференцированные пирамидоподобные нейроны (57,1 %) – в РАС, густоветвистые корковые средних размеров, подкорковые и переходные между ними формы нейронов (67,7 %) – в СОр.
В палеокортикальных структурах выражена слоистость, между слоем плотноупакованных клеток и глубоким клеточным слоем присутствует прослойка белого вещества - сетчатый слой, в их составе преобладают асимметричные поляризованные нейроны, характеризующиеся упорядоченностью в пространстве. В межуточных формациях клетки располагаются диффузно, здесь не обнаружены зоны переплетения отростков, входящие в их состав нейроны характеризуются большим разнообразием основных и переходных форм (рис.).
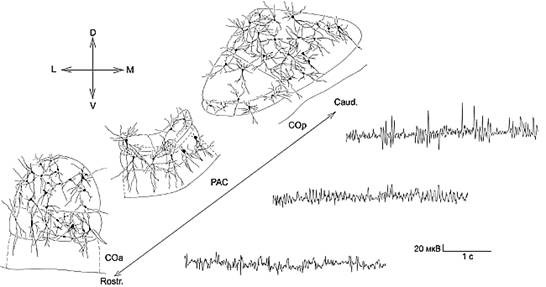
Рис. Нейронная организация СОа, РАС, СОр и характерный для каждого отдела паттерн ЭЭГ (L–M, D–V, Rostr.–Caud. – оси мозга).
Опираясь на существующее в литературе мнение о важном значении дендритов в функциональной организации нервной системы [2] и данные о том, что для определенного вида нейронов, в частности, мультиполярных и веретеновидных клеток, характерен определенный паттерн отводимой от них фоновой импульсной активности [14], мы предположили, что выявленные нами особенности нейронной организации СОа, РАС, СОр, а именно преобладание определенных форм нейронов, пространственное их расположение, степень разветвленности и протяженности отростков, а также разобщенность трех отделов в пространстве, не могут не сказаться на физиологических особенностях отделов кортикального ядра.
Высказанное предположение получило подтверждение в ходе электрофизиологического исследования. В табл. 1 приведены средние значения относительной спектральной плотности колебаний в ![]() -,
-, ![]() -,
-, ![]() -,
-, ![]() 1- и
1- и ![]() 2-диапазонах на ЭЭГ СОа, РАС и СОр.
2-диапазонах на ЭЭГ СОа, РАС и СОр.
Таблица 1. Результаты спектрального анализа фоновой ЭЭГ трех отделов кортикального ядра.
| Диапазон | Относительная спектральная плотность, Х |
||
| СОа | РАС | СОр | |
| 42,61 |
36,46 |
28,41 |
|
| 19,91 |
21,61 |
21,16 |
|
| 14,68 |
16,39 |
17,77 |
|
| 13,14 |
12,47 |
16,47 |
|
| 9,76 |
13,06 |
16,20 |
|
Примечание: Х - среднее значение, Sx - стандартная ошибка среднего.
Проведенный далее дисперсионный анализ влияния локализации регистрирующего электрода на спектральный состав ЭЭГ показал наличие статистически значимого действия этого фактора на относительную спектральную плотность колебаний в ![]() -,
-, ![]() -,
-, ![]() 1-,
1-, ![]() 2-диапазонах (табл. 2). Выявлено достоверное снижение представленности
2-диапазонах (табл. 2). Выявлено достоверное снижение представленности ![]() -ритма и увеличение относительной спектральной плотности колебаний в
-ритма и увеличение относительной спектральной плотности колебаний в ![]() -,
-, ![]() 1- и
1- и ![]() 2-диапазонах в ряду СОа–РАС–СОр.
2-диапазонах в ряду СОа–РАС–СОр.
Таблица 2. Влияние фактора локализации регистрирующего электрода на спектральный состав фоновой ЭЭГ кортикального ядра.
| Диапазон | F | p | |
| 64,40 | 0,0000 | 88,34 | |
| 0,65 | 0,5381 | 7,08 | |
| 5,54 | 0,0142 | 39,47 | |
| 10,11 | 0,0013 | 54,32 | |
| 15,88 | 0,0001 | 65,13 |
Примечание: F - коэффициент Фишера; ![]() 2 - коэффициент Ван-дер-Вардена.
2 - коэффициент Ван-дер-Вардена.
Переднее кортикальное ядро и заднее кортикальное ядро как межуточные формации, несмотря на различное рострокаудальное положение, имеют ряд общих черт, а именно: разнообразие представленных паттернов ЭЭГ и достаточно четкую обособленность отдельных ритмических компонент. В отличие от переднего и заднего отделов, для электрической активности РАС характерна дизритмичность и полифазность колебаний, при этом выделение превалирующего ритма представляется затруднительным. В то же время ЭЭГ РАС отличается большей однородностью и устойчивостью описанного типа активности.
Преобладание поляризованных упорядоченных в пространстве нейронов, наличие обширных проекционных входов, обеспечивающих поступление информации от первичных обонятельных областей (основной и добавочной обонятельных луковиц), как прямых, так и опосредованных через СОа и СОр, а также прямые двусторонние связи с пириформной корой, обуславливают тесную вовлеченность РАС в процессы интеграции и обработки сенсорной информации (главным образом обонятельной). Полученные нами данные могут свидетельствовать о функциональной роли периамигдалярной коры как важного ассоциативного центра миндалевидного комплекса.
Суммируя вышесказанное, необходимо подчеркнуть, что выявленные нами структурные особенности СОа, РАС и СОр (принадлежность к корковым и межуточным формациям, различия в нейронной организации) нашли отражение в проявлении их функциональной активности. Различия в цитоархитектонике и нейронной организации, а также электрофизиологические данные говорят о различном функциональном значении СОа, РАС и СОр в интегративных и регуляторных влияниях миндалевидного комплекса, а также о существовании рострокаудального градиента в функциональной организации кортикального ядра, что подтверждает концепцию о наличии рострокаудального градиента в его структурно-функциональной организации [1]. При этом каждый из трех отделов является пространственно организованной совокупностью функционально и структурно взаимосвязанных пирамидных и непирамидных нейронов, что предполагает различную роль отделов кортикального ядра в интегративных влияниях и функциональной регуляции деятельности мозга.
Исследования поддержаны грантом Министерства образования РФ.
Список литературы
Акмаев И.Г., Калимуллина Л.Б. Миндалевидный комплекс: функциональная морфология и нейроэндокринология. М.: Наука, 1993.
Scalia F., Winans S.S. The differential projection of the olfactory bulb and accessory olfactory bulb in mammals // J. Comp. Neurol. 1975. V. 161. № 1. P. 31-56.
Krettek J., Price J. A description of the amygdaloid complex in the rat and cat with observation on intra-amygdaloid axonal connections // J. Comp. Neurol. 1978. V. 178. P. 255-280.
Майковский Е. Патогенез эпилептического очага у человека // В сб.: “Актуальные вопросы стереонейрохирургии эпилепсии” СПб.: РНХИ им. А.Л. Поленова, 1996. C. 74-81.
Чепурнов С.А., Чепурнова Н.Е. Миндалевидный комплекс мозга. М.: Изд-во МГУ, 1981.
Чепурнов С.А., Чепурнова Н.Е., Маджид Б. Шейх, Нозмо Ф. Мукииби. Миндалина в экспериментальной эпилепсии // Актуальные вопросы стереонейрохирургии эпилепсии. СПб.: РНХИ им. А.Л.Поленова. 1996. С. 112-121.
Cain D.P. Kindling and the Amygdala / The Amygdala: Neurobiological Aspects of Emotion, Memory, and Mental Dysfunction. N.Y.: Wiley-Liss, 1992. P. 539-560.
Pitk![]() nen A., Tuunanen J., K
nen A., Tuunanen J., K![]() lvi
lvi![]() inen R., Partanen K., Salmaper
inen R., Partanen K., Salmaper![]() T. Amygdala damage in experimental and human temporal lobe epilepsy // Epilepsy Res. 1998. V. 32. P. 233-253.
T. Amygdala damage in experimental and human temporal lobe epilepsy // Epilepsy Res. 1998. V. 32. P. 233-253.
Леонтович Т.А. Нейронная организация подкорковых образований переднего мозга. М.: Медицина, 1978.
Поляков Г.И. Основы систематики нейронов новой коры большого мозга человека. М.: Медицина, 1973.
Paxinos G., Watson C. The Rat Brain in Stereotaxic Coordinates. San Diego: Acad. Press, 1998.
Заварзин А.А. Труды по теории параллелизма и эволюционная динамика тканей. Л.: Наука, 1986.
Pigache R.M. The Anatomy of “paleocortex”: A critical review // Adv. Anat., Embryol. and Cell Biol. 1970. V. 43. № 6. P. 1-62.
Бабминдра В.П., Брагина Т.А. Структурные основы межнейронной интеграции. Л.: Наука, 1982.
Reyes A., Lujan R., Rozov A., Burnashev N., Somogyi P., Sakmann B. Target-cell-specific facilitation and depression in neocortical circuits // Nature Neurosci. 1998. V. 1. № 4. P. 279-285.